

Introduction
Infectious hypodermal and hematopoietic necrosis virus (IHHNV) was first recognized in 1981 in cultured blue shrimp (Penaeus stylirostris) from South and Central America,1 and soon after it appeared in other penaeid shrimps of America and Asia.2 IHHNV is the smallest virus (20–22 nm) among shrimp pathogenic viruses, and it is non-enveloped and icosahedral. IHHNV has a 1.40 gm/mL density in CsCl (Cesium chloride) gradient suspension and a linear single-stranded DNA of 4.1 kb size.3 Based on the physicochemical and ultrastructural properties and partial characterization of viral proteins, IHHNV is tentatively assigned to the Parvoviridae family.4 A bent rostrum, wrinkled legs, rough cuticles,5 stunted growth,6 and shrunken antennae are typical symptoms of IHHNV. In addition, IHHNV does not cause mass mortality but a high level of deformities causing runt deformity syndrome in Penaeus monodon and Litopenaeus vannamei.7 IHHNV-diseased L. vannamei exhibit cuticular malformations of the rostrum and reduce growth rates associated with a chronic disease, which may be due to an abnormal glycolysis disorder known as the Warburg effect.8 The histological characterization of IHHNV infection relies on the demonstration of cowdry type “A” inclusion bodies in the tissues of ectodermal (gills, cuticular epidermis, nerve cord, and ganglia) and mesodermal (hematopoietic organ, antennal gland, gonads, lymphoid organ, connective tissue, and striated muscle) origin.9 Lightner and Redman10 noted that 10%–50% of the cultured shrimp population was affected, which led to huge economic losses in the shrimp industry. In farms, shrimp infection may result from horizontal transmission through ingestion of dead and moribund shrimps, and vertical transmission has been detected through infected ovarian tissues and ovocytes.11 Apart from these morphological alterations, IHHNV-infected shrimps in our study also showed hyperplasia. Reports of tumor-like growth are rare in crustaceans, with only six significant cases documented. Spark et al.12 reported a tumor-like papilliform growth in brown shrimp (Penaeus aztecus). Overrstreet and Van Devender13 described a non-malignant tumor known as hamartoma in commercial shrimps, which was induced by environmental pollution. Tumor-like hernia in blue shrimp (Penaeus stylirostris) was reported by Lightner et al.14 Sparks and Morado15 observed a carcinoma-like neoplasm in an adult female red king crab (Paralithodes camtschatica). Lymphatic neoplasm from hematopoietic tissue was observed in L. vannamei by Lightner and Brock16 and an unusual, large, blue tumor-like growth reported in lobster Homarus americanus was described by Shields and Small.17 At present, there is no known treatment to control IHHNV, but viruses could be prevented through holistic approaches.18 Chemotherapy, vaccination, and good management practices (GMP) can prevent losses caused by diseases.19 Despite several previous reports in India regrading IHHNV infection in Penaeus monodon, we reported IHHNV infection in L. vannamei cultured in high densities. IHHNV is an emerging threat to the Indian shrimp culture industry. This study describes the morphological and cellular lesions caused by IHHNV infection by viral load in the gill cells of the infected shrimps and confirmed the identity of IHHNV by polymerase chain reaction (PCR).
Materials and Methods
Shrimp collection
Shrimp sampling was done from January to June 2015 in different locations along the Andhra Pradesh and Tamil Nadu shrimp farms in India. A cast net was used for the collection of shrimps from grow-out ponds. 34 semi-intensive ponds were selected based on geographical diversity, accessibility, and shrimp farming activity. The ponds ranged in size from 0.5 to 2 hectares. From each pond, 250–280 shrimps were examined for the presence of IHHNV. Infected shrimps were separated based on morphologically distinct characteristics unique to IHHNV, such as bent rostrum, stunted growth, shortened antennae, and shrunken eyes. The length and weight of the shrimps were measured using a digital vernier caliper and a precision balance, and morphological deformities were also noted. The normal and suspected IHHNV-infected shrimps were segregated and preserved for further analysis.
Histopathology
Normal and infected shrimps were aseptically dissected, and hepatopancreas tissue, muscle, and tumor-like lesions were preserved in Davidson’s fixative for 48 h. Then, the tissue samples were transferred to 70% ethanol and processed using the method described by Bell and Lightner.20 The tissues were dehydrated, embedded in paraffin wax, and sectioned at 3 µm using a microtome (YORCO, YSI 115). Sections were stained with hematoxylin and eosin and subsequently examined under a phase-contrast microscope (Nikon, ECLIPSE TS100LED-F MV). In all studies, normal and diseased shrimps were examined in triplicate.
Stereomicroscopic study
The antennae of normal and diseased shrimps were dissected using a sterile surgical blade. Both antennae were submerged in separate petri plates with water and observed under a stereomicroscope at different magnifications (Magnus MZR-TR). Stereomicroscopic observations were conducted at magnifications ranging from 10x to 40x, allowing detailed examination of the antennal segments and other morphological features.
Scanning electron microscopic (SEM) study
The primary antennae of both normal and diseased shrimps were preserved in 3% glutaraldehyde and dehydrated using a graded ethanol series (30%, 50%, 70%, 90%, and 100%). Following dehydration, the samples were dried using a critical point drier, and then sputter-coated with gold. The antennae were subsequently observed under a JEOL JSM 5610LV scanning electron microscope operating at 15 kV. SEM imaging was performed at varying magnifications to capture detailed morphological features.
Transmission electron microscopic (TEM) study
For electron microscopy, tissues were fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4) for 2 hours at room temperature and rinsed in the same buffer. The tissues were post-fixed for 1 hour in 1% osmium tetroxide in 0.1 M sodium cacodylate buffer and then washed in three changes of the same buffer. Following this, the samples were dehydrated through a graded acetone series and embedded in Agar 100 epoxy (Agar Scientific, Agar 100 pre-mix kit medium). The samples were polymerized overnight at 60°C in an oven. Semi-thin sections (1–2 µm) were stained with Toluidine Blue for light microscopy to identify suitable target areas. Ultrathin sections (70–90 nm) of these areas were mounted on uncoated copper grids and stained with 2% aqueous uranyl acetate and Reynolds’ lead citrate.21 The grids were examined using a JEOL JEM 1210 transmission electron microscope.
Molecular detection of IHHNV
Nucleic acid extractionPleopods and gills were used for DNA extraction to screen the systemic pathogen of IHHNV. The total DNA from the samples was isolated through DTAB-CTAB extraction using the IQ2000 Detection and Prevention System, GeneReach, Taiwan, in accordance with the manufacturer’s instructions. The quality of the isolated nucleic acids was confirmed using Bio Photometer Plus (Eppendorf, Hamburg, Germany).
Genomic identification of pathogen by PCR
Viral DNA was isolated from shrimp gill using a “high-purity DNA template preparation kit” (Roche Biosciences). The isolated DNA was subjected to PCR amplification using the specific primer. The 309F TCCAACACTTAGTCAAAACCAA and 309R TGTCTGCTACGATGATTA TCCA primers were used to amplify a partial region of the IHHNV non-structural gene. PCR (25 μL) was conducted using 100 ng of DNA and the EmeraldAmp GT PCR master mix (Takara Bio) in accordance with the manufacturer’s instructions. The thermal cycling conditions used for the PCR were as follows: 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s, and a final extension at 72 °C for 7 min.22 Following electrophoresis at 60 V for 30 min in 1× TBE buffer, the amplified genes were visualized under a UPV imaging system, and the amplified products were determined using a 100-bp ladder (GeneDirex, India).
Results
Sample collection and gross observation
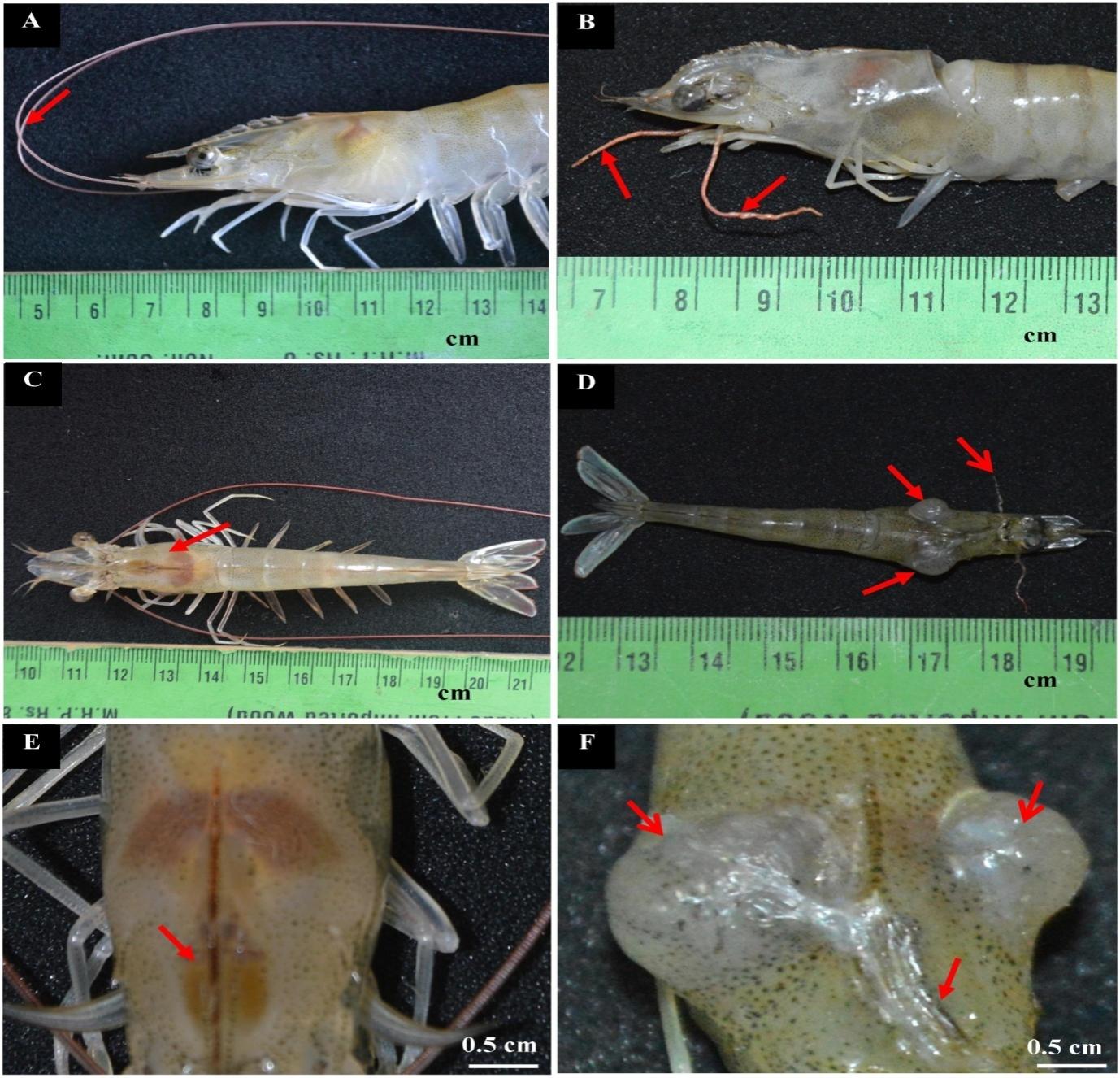
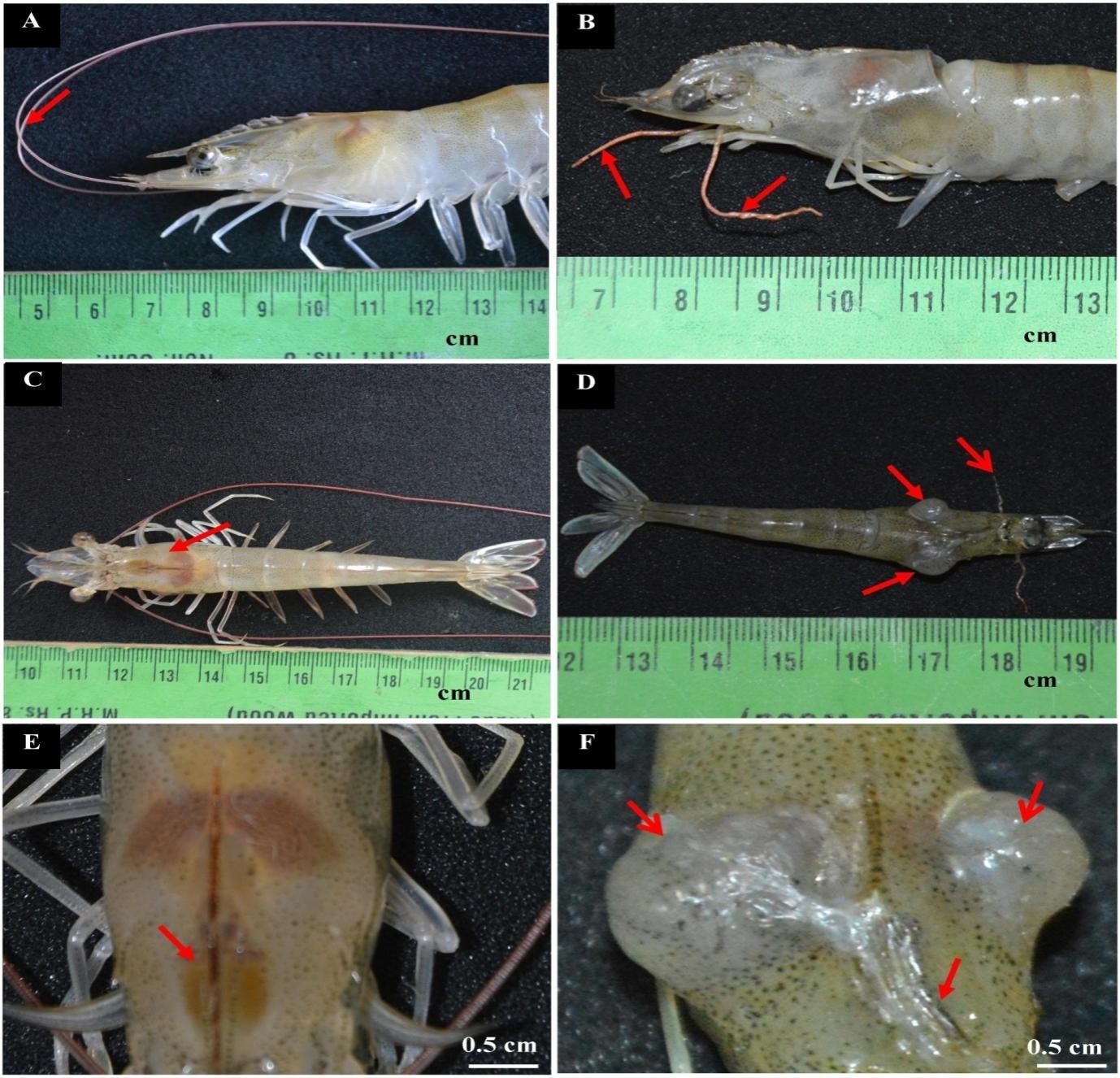
A total of 34 semi-intensive ponds were tested, and only one pond was reported to have IHHNV-diseased shrimps, which is situated in Kanuparthi village, Andhra Pradesh (Lat 15° 36′ 19.2132″ N, Long 80° 14′ 4.4556″ E). In addition, 43.26% of shrimps infected with IHHNV showed reduced body size with morphological aberrations. Normal shrimp length and weight ranged between 12.5–14.6 cm (12.8±0.5) and 22.2–25.5 g (23.4±2.04), respectively. The length of diseased shrimps ranged between 10.5 and 11.5 cm (10.8 ± 0.3), whereas the weight ranged between 9.37 and 14.12 g (10.8 ± 2.06 g). Normal shrimp were found to have straight rostrum, glassy cuticles, distinctly segmented abdomens, and spherical eyes (Figs. 1A, C, and E). The gross alterations observed in infected shrimps include bent and shortened rostrum (Fig. 1B), deformed abdomen with opaque abdominal musculature, ring-shaped deformed abdominal segment (Fig. 1D), and shrunken eyes (Fig. 1F). Lengthy and equally segmented antennae were observed in normal shrimp (Fig. 2A), but in IHHNV-diseased shrimp, the antennae were shortened (Fig. 2B). The carapace of normal shrimp was intact (Fig. 2C), whereas two tumor-like growths from the ventrolateral junction of the cephalothorax and abdomen were found on both sides of the infected shrimp carapace (Fig. 2D). The gastrofrontal sulcus was straight (Fig. 2E). Hyperplasia was fungating in shape and whitish in color. The growth on the right side of the carapace was larger (9 mm) than that on the left (5 mm). In the case of infected shrimp, the gastrofrontal sulcus was bent (Fig. 2F).

Stereomicroscopic study
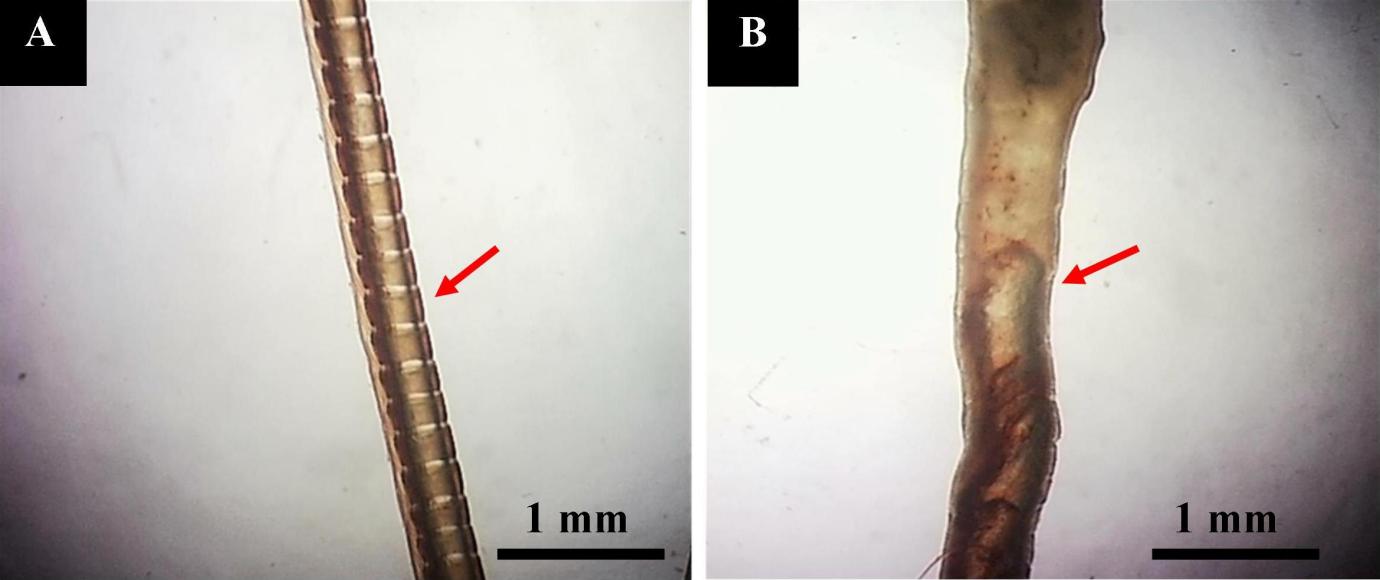
Stereomicroscopic observation shows that the normal antennae are segmented, and the entire organ was brown (Fig. 3A). However, the antennal segments of infected shrimp were fused, often indistinct, and irregular with brown and white colors scattered on the entire surface of the antenna (Fig. 3B).

Histopathological investigation
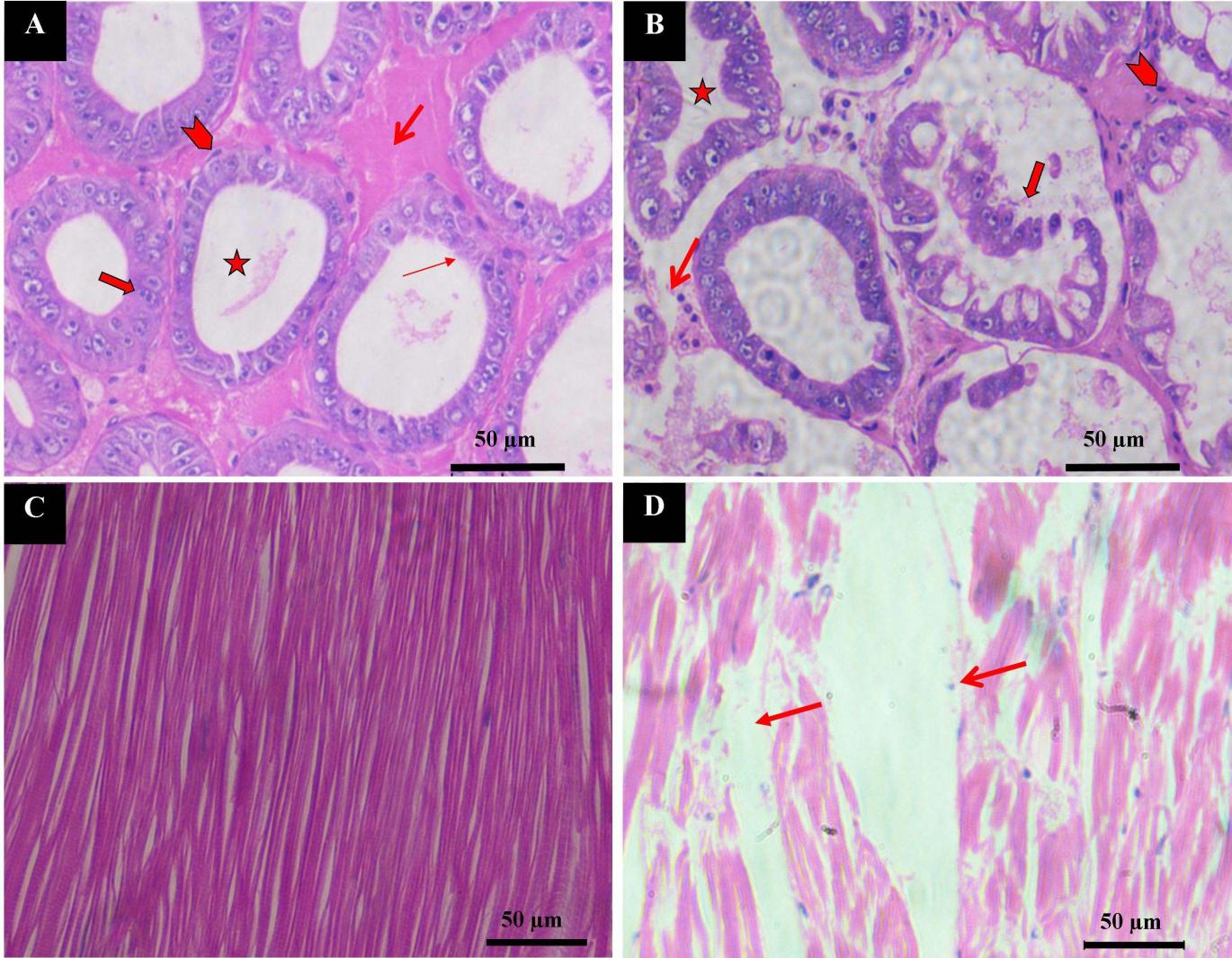
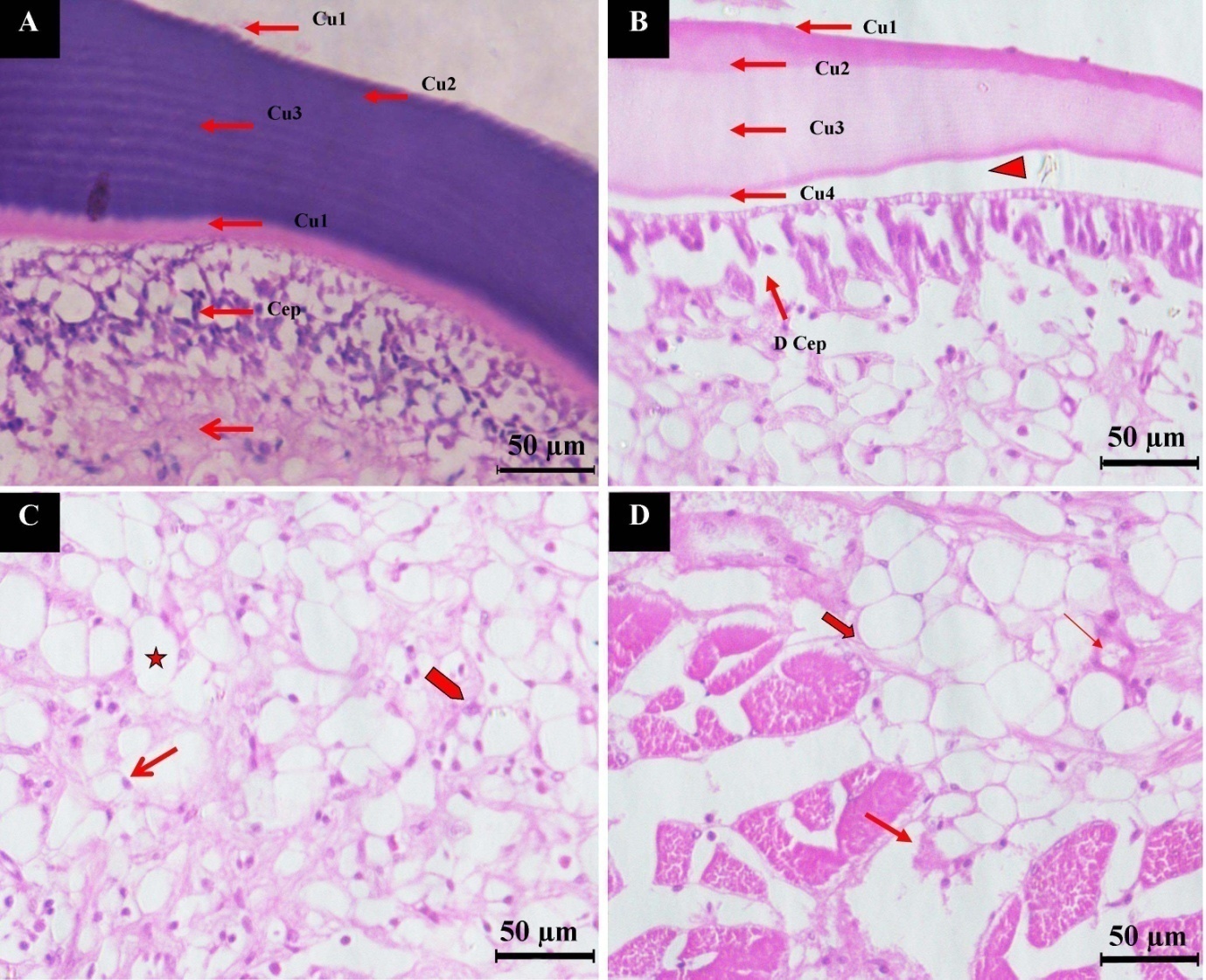
During the histological investigation, normal hepatopancreas tissue showed a large, ovoid lumen (35–70 nm) and a continuous microvillus brush border. Normal hepatopancreatic tubules were surrounded by connective tissue and myoepithelial cells, which were undergoing division (Fig. 4A). The hepatopancreatic tubule lumen was shrunken, and the epithelium had detached from the tubules and perhaps been missing in IHHNV-infected shrimps. The connective tissue between adjacent tubules was greatly diminished in size and degraded with empty spaces (Fig. 4B). The cross-section of normal muscle shows well-demarcated long cylindrical, unbranched striated muscle fibers (Fig. 4C). Figure 4D shows muscle from an infected shrimp, which was poorly defined with striated muscle bundles and scattered nuclei. A greater amount of space between muscle bundles was evident. The histological section of normal carapace shows all four sublayers (epicuticle Cu1, exocuticle Cu2, endocuticle Cu3, and cuticular membranous Cu4). A well-arranged cuticular epithelial layer (Cep) was attached to the cuticular membranous layer (Cu4) of the cuticle with denser connective tissue below. A normal cuticle is blue after staining, which indicates the presence of calcium salt (Fig. 5A). The cross-section of hyperplasia shows the four layers of the cuticle with modifications. The first layer, epicuticle (Cu1), was thin and attached to the second exocuticle (Cu2) sublayer. The exocuticle (Cu3) was wider than the epicuticle. The third sublayer (endocuticle) is the widest layer among the cuticle layers, which is attached to the cuticular membranous (Cu4) layer, known as the uncalcified layer. These layers were not parallel, and a gap was observed between the cuticle and cuticular epidermal layer (Cep). This cuticle was shrunken and merged together, and the damaged cuticular epidermal layer (D Cep) was visible. The cuticle of hyperplasia was pale pink, which indicates loss of calcium salt (Fig. 5B). Cells can also be seen in the cross-section of hyperplasia, which shows big vacuoles. In such cells, nuclei were present in the peripheral region, and some cells were under pyknosis (Fig. 5C) and karyorrhexis state. Degrading muscle fibers were also intact with hyperplasia cells (Fig. 5D).


SEM study
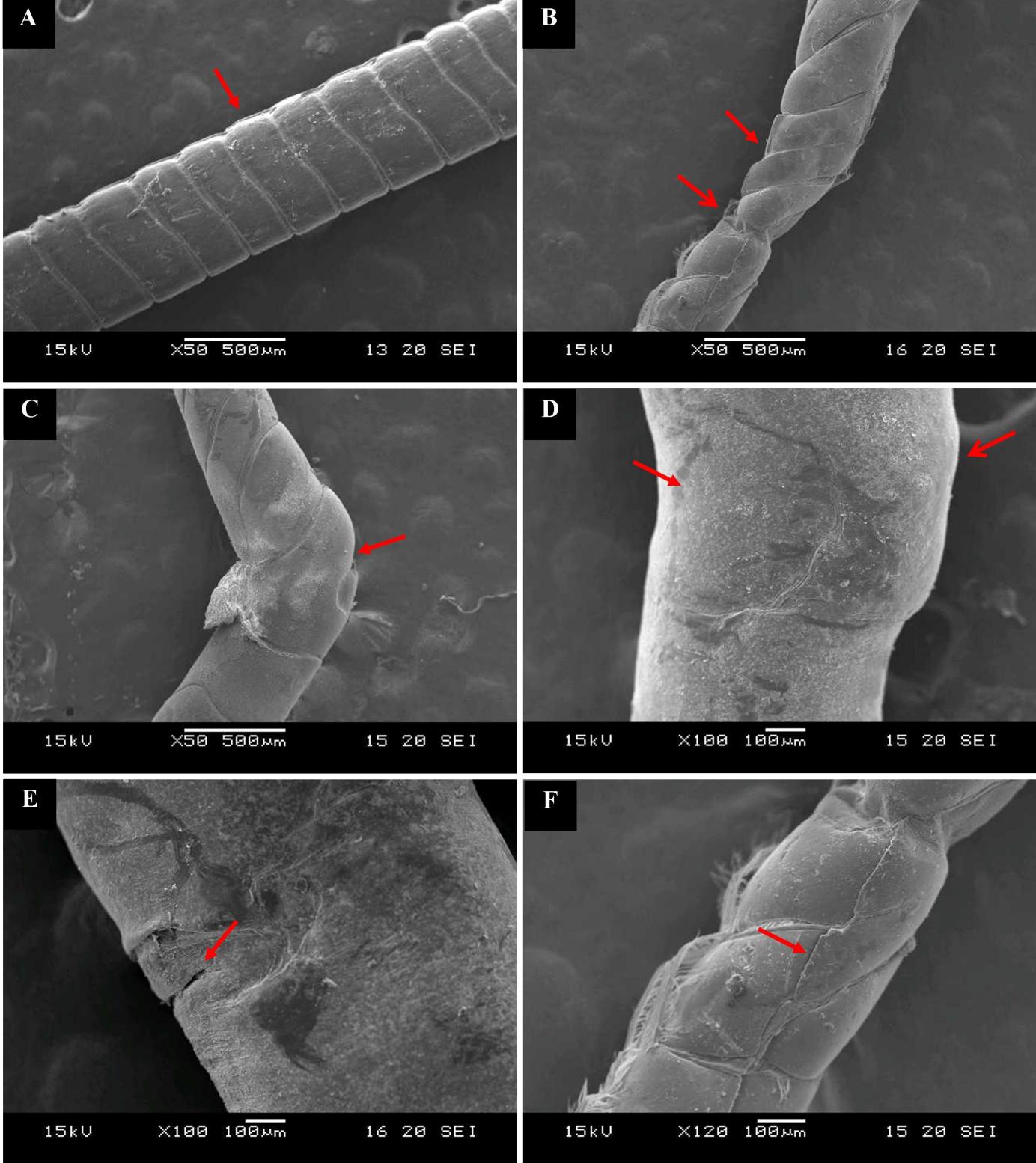
The SEM images show that the antennae of normal shrimps are evenly segmented with a smooth surface and cylindrical shape (Fig. 6A), whereas the IHHNV-infected shrimp antennal segments were unequal, twisted, shrunken (Fig. 6B), and bent (Fig. 6C). In some areas, the antenna was swollen, dented (Fig. 6D), and cracked in horizontal and (Fig. 6E & F) vertical pattern.

TEM study
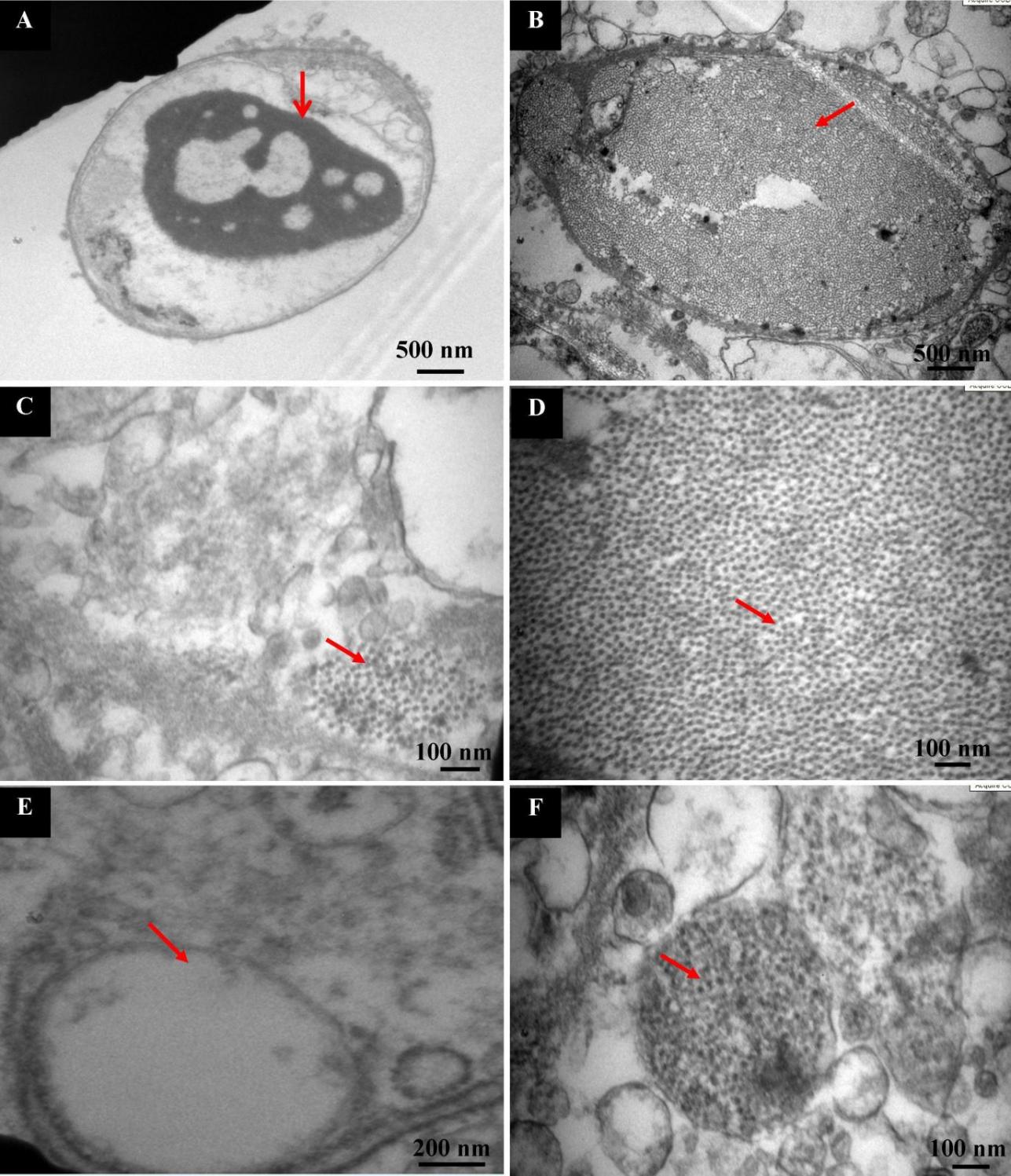
TEM observation of the gill cells of normal shrimps shows intact nuclei with dense centralized chromatin without any viral particles (Fig. 7A). On the contrary, in infected gill cells, the nuclei had lost distinctive heterochromatin, and they were filled with viral particles (Fig. 7B). The viral particles were also present in the cytoplasm of infected cells in a scattered manner (Fig. 7C), and the viral particles stipate (crystal-like arrangement) to one another under the magnified observation (Fig. 7D). The vacuoles of the normal cell were empty (Fig. 7E), but in the case of IHHNV-diseased shrimp, the gill cells showed viral particles in vacuoles too (Fig. 7F). TEM observation of normal shrimp muscle revealed striated fibers with well-arranged A band and I band. The H zone, which consists of the M line, was also present in the middle of the A band. The Z line was present in the middle of the I band (Fig. 8A). The IHHNV-infected muscle showed areas where myofibrils were split, and the oval-shaped nucleus was present, in which chromatin was in scattered form (Fig. 8B). In normal muscle, all myofibrils were intact and clearly visible in higher magnification (Fig. 8C), whereas the infected muscle showed a gap between the fibrils (Fig. 8D).


Molecular diagnosis
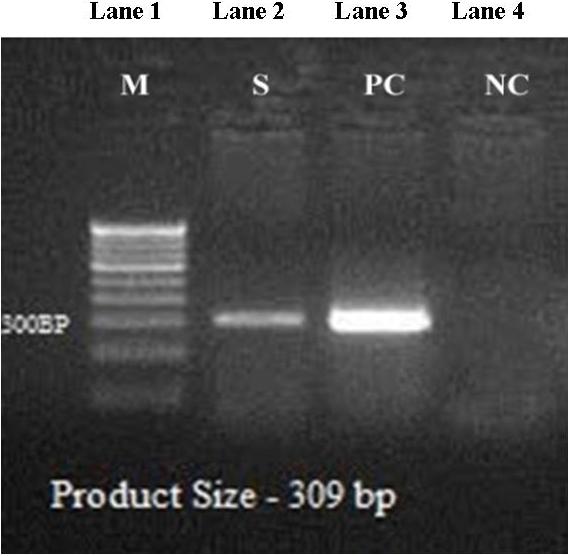
Under the optimized PCR condition, a single viral DNA product of a predictable size of 309 bp was detected. The amplified DNA fragment of infected shrimp was compared with positive control, which showed the same molecular weight. Normal shrimp, which was taken as the negative control, did not show any DNA band (Fig. 9).

Discussion
The infectious hypodermal and hematopoietic necrosis virus (IHHNV) is known to cause abnormalities in the development and growth in shrimp, and infections with the Litopenaeus vannamei virus have been linked to 10% to 50% of economic losses.10 In Litopenaeus vannamei, a persistent infection not only inhibits growth but also results in rostrum, antenna, thoracic, and abdominal cuticular abnormalities. Runt deformation syndrome (RDS) is the term used to describe these combined clinical symptoms.6 In the present study, the gross clinical, histological, microscopic, and molecular results were consistent with IHHNV infection, contributing to morbidity with stunted growth of L. vannamei. In gross observation, the shortened rostrum and short twisted antennae are the key characteristic features of IHHNV infection, but other symptoms such as hyperplasia in cephalothorax, rough cuticle, and deformed abdominal segment are the additional characteristic features of IHHNV infection of L. vannamei. Reduced growth rates and a variety of cuticular deformities affecting the rostrum (‘bent rostrum’), antennae, and other thoracic and abdominal areas of the exoskeleton were also reported previously.6,9,23 Hematopoiesis is the process in which various types of cells are produced from the stem cells. IHHNV may alter the production of hypodermal cells, and it may be drastically reduced by necrosis, which may be the reason for the deformities. In the larval stages, IHHNV-infected shrimp may not exhibit the conspicuous symptoms observed in adult larvae, so molecular investigation is necessary to confirm the infection. IHHNV infection is acute in small shrimps rather than in larger ones. Kalagayan et al.6 observed that small shrimp had 100% infection, whereas the large cohorts had only 70% infection. Young shrimp are more vulnerable than the adult ones. Dewangan et al.24 reported that co-infection with WSSV and IHHNV resulted in shrimp mortality, whereas shrimp infected only with IHHNV survived. The present study observed that IHHNV infection accounted for 43.26% of the shrimp population in the pond of Andhra Pradesh. A screening for infectious hypodermal and hematopoietic necrosis virus (IHHNV) was conducted on 104 wild-caught Litopenaeus vannamei broodstock that had been taken off the Pacific coast of Panama in March 2000. In 28% of the 104 hemolymph samples examined, the dot blot assay findings showed the presence of IHHNV.25 Vertical transmission may be crucial for the increased IHHNV infection in domesticated shrimps from one generation to another generation.26 In India, IHHNV was first reported in cultured Penaeus monodon from Chidambaram, Tamil Nadu.27 Moreover, broodstock of L. vannamei has been imported from other countries to produce offspring in shrimp hatcheries. These broodstock are considered specific pathogen-free (SPF). This strategy would help prevent vertical transformation. The offspring of SPF broodstock will be free from viruses, including IHHNV. These farm-reared shrimps might be affected by horizontal transmission. IHHNV infection in L. vannamei is more serious than that in P. monodon28 because L. vannamei is the dominant candidate species in Indian shrimp culture.
Necroses of hepatopancreatic cells led to the disintegration of the above-mentioned cells and thus affect the physiological function of the digestion and detoxification process. The infection also degraded the muscle cells and infiltration of the nucleus, which led to the loss of body flexibility and thus affected the locomotion of shrimp. Pazir et al.29 also found cellular degeneration, severe nuclear hypertrophy and margination of chromatin in tissues, especially in the hepatopancreas, cuticular epithelium, gills, connective tissues, hematopoietic tissues, and digestive epithelium. The hyperplasia in the IHHNV-infected shrimps is new documentation in the present investigation. The histology of hyperplasia revealed the presence of vacuolated cells and an atypical hyperchromatic nucleus. The first line of defense against microbial invasion is a robust exoskeleton made of chitin, calcium salts, and melanin, as well as the midgut’s peritrophic membrane.30,31 Carbohydrates and calcium salt are present in all four layers of the normal cuticle, which can be stained by hematoxylin. The normal cuticle revealed the presence of calcium salt (purple color), but the cuticle of hyperplasia had lost the calcium salt. TEM observation suggested that the virus would have used the host cell machinery for their replication and would have occupied not only the entire nucleus but also the vacuoles. This condition could interfere with all the physiological and biochemical functions of the infected tissue, which may lead to the uneven growth of tissue. Antennae are the most important sensory organs in shrimp, playing a crucial role in environmental sensing and recognizing the opposite sex. Furthermore, this abnormal condition may lead to difficulty in locating food, thereby leading to the reduced growth of shrimp. In addition, the hepatopancreas of infected shrimp shows major disruption of the cells involved in digestion, absorption, and excretion.32 Thus, necroses of hepatopancreatic cells may further affect the physiological function of digestion and detoxification. The IHHN-induced degradation of the muscle fibers led to the loss of voluntary and involuntary movements of shrimp and internal organs. The reduced cuticle thickness of the diseased shrimps would allow for secondary infection by bacteria, fungi, protozoa, and other viruses. This study demonstrates the distinctive features between IHHNV infections from other viral diseases of shrimps. The continuous assessment of IHHNV infection is necessary before the outbreak of the disease in the Indian shrimp industry.
Acknowledgments
The authors would like to thank the Dean and Director of the Center of Advanced Study in Marine Biology, Faculty of Marine Sciences, Annamalai University, for providing facilities during the study period. The authors are also thankful to the Department of Science and Technology (DST-SEED), Government of India, Project No. SEED/TITLE/2019/96©, and Initial Scientific Research Fund of Young Teachers (2023HNUAHEDF009,2022HNUAHEDF023), Scientific and Technological Project of Henan Province (242102110072,242102111013,242102111024), the Special Fund for Henan Agriculture Research System (HARS-22-16-Z1)for financial support in carrying out the said task. We are also thankful to shrimp farmers and farm technicians for extending their help during the sample collection from their grow-out farms. We would like to thank Ms. A. Bascal Rita, C. Gunasekaran, and CMC Vellore for transmission electron microscopic studies. We would also like to thank the Central Instrumentation Facility (CIF) of Annamalai University for SEM analysis.
Authors’ Contribution
Conceptualization: Caiyuan Zhao (Lead). Writing – original draft: Caiyuan Zhao (Lead). Formal Analysis: Naresh Kumar Dewangan (Equal), Ramakrishna Rajkumar Singh (Equal). Investigation: Naresh Kumar Dewangan (Equal), Ramakrishna Rajkumar Singh (Equal). Supervision: Ayyaru Gopalakrishnan (Lead). Methodology: Savva Sravani (Equal), Anisha Shafni John (Equal). Writing – review & editing: Wenyan Xu (Equal), Guojun Guo (Equal), Silei Xia (Equal), Sonu Tiwari (Equal), Xudong Li, Gaixiao Qin (Equal), Jianan Hou (Equal).
Competing of Interest – COPE
The authors declare no competing interests
Ethical Conduct Approval – IACUC
Manuscript does not need an ethical approval.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.