Introduction
Cellulose is an important component of plant cell wall and it is present in a relatively large content in raw materials of vegetable-based feeds. Herbivorous grass carp (Ctenopharyngodon idellus) and blunt snout bream (Megalobrama amblycephala) digest cellulose from aquatic plants depending on the cellulose produced by the cellulolytic bacterial community in their intestinal tracts.1–3 Cellulase is the generic term for a group of complex enzyme systems. It is mainly divided into three types based on function: 1) β-1,4-endoglucanases, which act on the non-crystalline region inside the cellulose, hydrolyses β-1,4 glucosidic bonds randomly, cuts long-chain cellulose molecules and produces abundant cellulose with non-reduced terminal micromolecules; 2) cellobiohydrolases, which act on the terminal of cellulose linear molecules, hydrolyses β-1,4 glucosidic bond, and cleaves cellobiose molecules. Hence, it is also known as cellobiose hydrolase; 3) β-glucosidases, which generally hydrolyses cellobiose or soluble cellodextrin into glucose molecules.4,5
Several microorganisms form complex communities in animal digestive tracts, resulting in a diverse ecosystem.6 Intestinal microorganisms form a symbiotic relationship with the host.7 They play an important role in nutrient absorption, growth, metabolism, host resistance and immune response to disease.8–11 Several cellulolytic bacteria have been separated successfully from the intestinal tracts of grass carp, including Aeromonas, Enterococcus, Bacillus, Enterobacterium, and Citrobacter sp.12–15
Red swamp crayfish (Procambarus clarkii) is an important aquaculture shellfish in China. Between 2007 and 2022, the aquaculture output of P. clarkii in China increased from 265,500 tons to 2.8907 million tons. In 2022, the commercial value of P. clarkii industry in China was about 458 billion yuan. As omnivores, P. clarkii prefers aquatic plants and their intestinal tracts degrade cellulose, which might be related to the endogenous cellulose16 and the presence of cellulose-degrading bacteria in the intestine. Feng et al.17 screened cellulolytic bacteria, including Bacillus velezensis and B. subtilis, from the intestinal tracts of P. clarkii via pure culture method using a carboxymethylcellulose medium. There are few studies evaluating the cellulase activities of P. clarkii by using microcrystalline cellulose and cellobiose medium. In the present study, cellulolytic bacteria which can produce multiple cellulase types were obtained by screening from the intestines of P. clarkii using carboxymethylcellulose, microcrystalline cellulose and cellobiose. It provides a theoretical reference to develop probiotics in the feed supplied to P. clarkii.
Materials and Methods
Experimental animals
Twelve healthy red swamp crayfish (20.32±0.95g) were obtained from a local farm in Changde, Hunan Province, China in June. The ratio of male to female was 1:1.
Preparation of medium
Three media were prepared as described by Li et al.2 The carboxymethyl-cellulose(CMC)-agar medium (1L) is composed of 10 g CMC, 4 g KH2PO4, 4 g Na2HPO4, 2 g tryptone, 0.2 g MgSO4·7H2O, 0.001g CaCl2, 0.001g FeSO4·7H2O, and 15 g agar powder. The microcrystalline cellulose (MC)-agar medium (1L) is composed of 10 g microcrystalline cellulose, 4 g KH2PO4, 4 g Na2HPO4, 2 g tryptone, 0.2 g MgSO4·7H2O, 0.001 g CaCl2, 0.001g FeSO4·7H20, and 15 g agar powder. The cellobiose (GC)-agar medium (1L) comprises 4 g esculoside, 1g ammonium ferric citrate, 10 g tryptone, 5 g yeast extract, 10 g NaCl, and 15 g agar powder. The tryptic soy agar (TSA) medium uses the TSA powder at a 40g /L mixing concentration.
Screening of cellulolytic bacteria
Intestinal tracts of 12 P. clarkii samples were extracted under aseptic condition. The intestinal tracts and sterile PBS (0.1 mol/L, pH 7.2) were homogenised at the ratio of 1:9. The intestinal-PBS mixture was obtained after homogenization. The mixture was processed via serial dilution:10-2, 10-3, 10-4, 10-5 and 10-6. Each dilution contained two preparations and was coated onto the TSA plate, followed by incubation at 28°C for 48 h. Several single colonies were selected for streaking on the TSA plate, and cultured for 48 h at 28°C. After purification three times, the bacteria were inoculated onto the CMC-agar medium, MC-agar medium and GC-agar medium via point inoculation. They were cultured for 48 h at 28°C, and then dyed with 5 mL 1 mg/mL Congo red for 10 min, followed by decolourisation with 1mol/L NaCl solution for 5 min. The bacteria with transparent hydrolytic ring represented cellulolytic bacteria. Next, the top 5 bacteria with better cellulose degradation capacity were screened based on the speed of the transparent hydrolytic ring and the size of the bacterial colony. The diameter of the hydrolytic ring surrounding the colony (D) and the colony diameter (d) on the plate were measured with a microscope. Each strain was measured 3 times and the D/d ratio was used as the index for preliminary determination of the degradation ability. A statistical analysis of D/d values was performed by using SPSS 20.0 software.
Molecular biological identification of strains
The colonies were cultivated in TSB at 28°C for 24 h, and harvested by centrifugation at 4000 g for 10 min. DNA was extracted using a bacterial genomic DNA extraction kit (Tsingke Biotechnology Co., Ltd,Beijing, China). The 16S rRNA gene sequence was amplified using primers 27F (5’-AGAGTTTGATCCTGGCTCAG-3’) and 1492R (5’-TACGGCTACCTTGTTACG ACTT-3’) according to the method described by Weisburg et al.18 Following the completion of the PCR reaction, the PCR products were detected via 1% agarose gel electrophoresis (AGE). The amplified DNA segments were sent to Tsingke Biotechnology Co., Ltd for DNA sequencing. Later, BLAST comparative analysis of sequences was carried out in the NCBI database (blast.ncbi.nlm.nih.gov) to determine the species relationships. The phylogenetic tree was constructed using the neighbor-joining (NJ) technique in MEGA7.0 software.
Results
Isolation and screening of intestinal cellulolytic bacteria in red swamp crayfish
A total of 12 single colonies on TSA plate were selected for analysis. They were separated and purified repeatedly on three cellulose plates. After treatment with Congo red, five strains produced hydrolytic rings. As shown in Table 1, the 5 cellulolytic bacterial strains were E, M, G, C and H on the CMC-agar medium ranging in size from big to small, E, C, M, G and H on the microcrystalline cellulose plate (Table 2), as well as H, G, C, M and E on the cellobiose plate (Table 3). The strain E presented the strongest degradation of cellulose in both CMC-agar medium and MC-agar medium. Although strain H had a weak ability to decompose cellulose directly, it exhibited the strongest capacity for the decomposition of cellobiose.
Strain C was a small and smooth yellowish white round colony. It contains an obvious bulge in the center, with smooth and wet edges. Strain E was a yellow oval colony, with obvious boundaries between the center and edges. The edges were extremely thin and it was thick in the center, with a rough surface. Strain G was a milk-white oval colony, with smooth and wet surfaces. It carried a yellowish and swollen center. Strain H was a light red large round colony, without obvious boundaries between the center and edges. The center was slightly thick and white, while the edges were light red. The surface was smooth and wet. Strain C was a light yellow round colony with moderate size and mild boundary sense at the edges. The external edges were white and the central bulge was yellow.
Identification of strains and construction of the phylogenetic tree
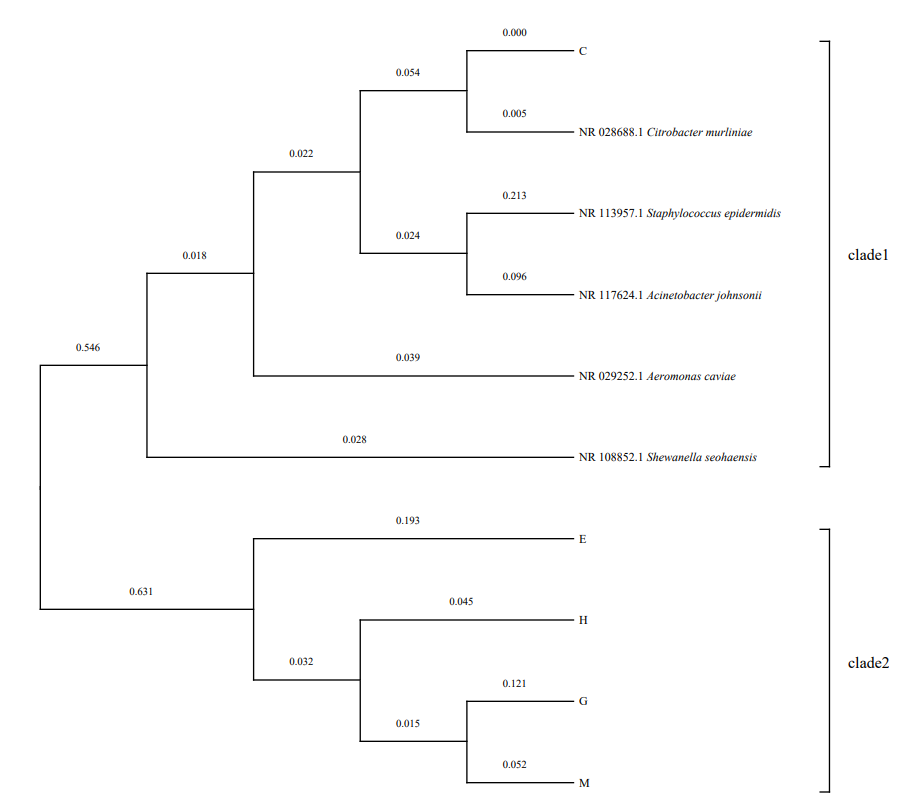
The 16S rDNA gene of the screened strains was amplified by PCR to obtain nearly 1500 bp. The 16S rDNA sequences (about 1500 bp) of the five screened cellulolytic bacteria were submitted to GenBank (Table 4). Similarity search was performed on NCBI database. The 16S rDNA sequence similarity between strain C and Citrobacter sp. reached 99.86%. The 16S rDNA sequence similarity between strain E and Staphylococcus sp. was 100%. The 16S rDNA sequence similarity between G and Acinetobacter johnsonii reached 99.71%.The 16S rDNA sequence similarity between strain H and Shewanella sp. was 99.79%. The 16S rDNA sequence similarity between strain M and Aeromonas caviae was 99.79%. The phylogenetic tree (Figure1) showed that strains H and M had a close genetic relationship and strain C was clustered independently, indicating that strain C had a distant genetic relationship with other four strains.

Discussion
Five cellulolytic bacteria were screened from the intestinal tract of P. clarkii via pure culture method in this experiment. According to the results of 16S rRNA molecular identification, the strains C, E, G, H, and M were Citrobacter sp., Staphylococcus sp., Acinetobacter johnsonii, Shewanella sp., and Aeromonas caviae, respectively. Feng et al.17 screened cellulolytic bacteria from the intestinal tracts of P. clarkii using CMC-agar medium, and were identified as B. velezensis and B. subtilis. The screened bacteria differed from those detected in this experiment, which might be attributed to differences in intestinal flora caused by different living environments.19
Deng et al.20 explored the intestinal flora of P. clarkii cultured in ponds via high-throughput sequencing and found that the dominant bacteria included predominantly Citrobacter sp., Hafnia enterococcus, Candidatus Bacilloplasma, Bacteroides sp., Clostridiaceae unclassified genus, and Shewanella. Feng et al.21 studied the intestinal flora of P. clarkii which were cultured in-door by high-throughput sequencing and found that the dominant bacteria were Candidatus Bacilloplasma, Bacteroides, Acinetobacter, Dysgonomonas, Tyzzerella3, Aeromonas, and Shewanella. Li et al.14 found that Aeromonas, Enterobacter, Enterococcus, Citrobacter sp. and Bacillus were the major cellulolytic bacteria in the intestines of grass carp. In this experiment, Citrobacter sp. and Shewanella sp. were the dominant culturable bacteria in the intestinal tract of P. clarkii and they play an important role in cellulose degradation of P. clarkii.
Huang22 found that Citrobacter sp. was the cellulolytic bacteria in the intestinal tract of Holotrichia parallela larvae, which was consistent with the results of the present study. He23 reported that Citrobacter sp. was a common nonpathogenic bacterium in animal intestinal bacteria. Deng et al.24 also found that Citrobacter sp. was a normal core flora in the intestinal tracts of P. clarkii. However, Chen et al.25 found that C. freundii was pathogenic in Micropterus salmoides. Liu et al.26 also found that C. freundii was a pathogenic bacterium associated with tail-rot disease of P. clarkii. Hence, the pathogenicity of Citrobacter sp. identified in this study will be further studied.
Aeromonas caviae is a pathogenic bacterial species infecting humans. It may adversely affect bile duct or urinary system and exhibits strong pathogenicity against Litopenaeus vannamei and Macrobrachium rosenbergii.27–29 Aeromonas caviae as cellulolytic bacterial species in animal intestines has yet to be reported. The pathogenicity of Aeromonas caviae in P. clarkii requires further investigation.
Staphylococcus acts as a lignocellulolytic bacterium infecting white ants.30 It is a cellulolytic bacterium infecting the intestinal tracts of Ornithophora auriculata larvae,31 which is consistent with our experimental findings. Staphylococci are a group of gram-positive cocci. S. aureus is one of the most important pathogenic bacteria, which cause global food poisoning. The screened staphylococcus in this experiment exhibits a completely different colonial morphology compared with S. aureus. Whether the screened Staphylococcus can be used as a probiotic for P. clarkii has yet to be investigated.
Acinetobacter johnsonii acted as a cellulolytic bacterial species in Coptotermes formosanus32 and degraded lignocellulose in the soils and straw compost.33 Acinetobacter johnsonii generally is not a pathogenic bacterium. Studies reported no human infection caused by Acinetobacter johnsonii.34 Acinetobacter johnsonii was not pathogenic in the intestinal tracts of grass carp.35 Hence, it has great potential for research and development as a probiotic.
Shewanella sp. was cellulolytic in the intestinal tract of P. clarkii in the present study. Shi36 also demonstrated that Shewanella sp. was cellulolytic in the intestinal tract of grass carp. Shewanella sp. is the dominant core intestinal bacteria in grass carp, and it can be used as a probiotic in aquatic animals. After feeding grass carp with Shewanella sp., the immunity was enhanced and the levels of total protein (TP), albumin, globulin and complement C3 level in serum were increased, accompanied by significant upregulation of IL-8, IL-1β, TNF-α and lysozyme-C.37 Hence, Shewanella sp. has great potential for use as a probiotic.
In summary, Aeromonas caviae is pathogenic to the human body and aquatic animals, and it cannot be used as a probiotic in P. clarkii. Citrobacter sp. and Staphylococcus sp. will be further evaluated and utilized for further research and development of probiotics. Acenatobacter johnsonii and Shewanella sp. have great potential for application as probiotics in forage for P. clarkia, suggesting the need for further studies. The present study can provide a theoretical basis for the development of probiotics in the feed of P. clarkia.
Acknowledgments
This work was supported by Hunan Provincial Department of Education Innovation Platform Project[19K064]; Hunan Provincial Aquatic Biological Resources Monitoring Station of Yuanjiang River and Lishui River[2130122]; Hunan Provincial Key Research and Development Program [2020NK2039];The Doctoral Start-up project of Hunan University of Arts and Science [18BSQD21]; Changde City science and Technology innovation Development Special Fund [2019S044]; Changde Special Key Project for Scientific and Technological Innovation and Development (No.[2021]67);The Innovation and Entrepreneurship Training Program for College Students (Hunan University of Arts and Science [2019] 20), The Innovation Team of Microbial Technology in Hunan University of Arts and Science [202026]. The authors declare no conflict of interest.
Authors’ Contribution
Conceptualization: Liye Shao (Equal), Pinhong Yang (Equal). Methodology: Liye Shao (Equal), Ronghua Wang (Equal). Data curation: Liye Shao (Equal), Xiangyan Qiu (Equal), Jiaxun Li (Equal), Junming Chen (Equal), Ronghua Wang (Equal). Investigation: Liye Shao (Equal), Ronghua Wang (Equal). Formal Analysis: Liye Shao (Equal), Xiangyan Qiu (Equal), Ronghua Wang (Equal). Writing – original draft: Liye Shao (Equal), Xiangyan Qiu (Equal). Writing – review & editing: Ronghua Wang (Equal), Pinhong Yang (Equal). Resources: Ronghua Wang (Equal). Funding acquisition: Pinhong Yang (Equal). Supervision: Pinhong Yang (Lead).