


Introduction
The global population is growing rapidly, leading to an increased demand for food.1 Ensuring that food provision is both safe and sustainable is of paramount importance. Over the past few decades, aquaculture has emerged as one of the fastest-growing animal food sources, owing to its rapid expansion and intensive development. It plays an increasingly important role in the global food supply.2–4 Infectious diseases caused by viruses, bacteria, and other pathogens are common occurrences in the aquaculture industry, resulting in substantial economic losses and significantly impacting its sustainable development.5 As aquaculture plays a vital role in global food security, understanding and managing the factors influencing fish health, such as gut microbiota dynamics during infections, contribute to ensuring a stable and secure food supply.
Diseases caused by bacterial infections, such as Aeromonas. spp, are a major cause of high mortality in farmed fish. These infections can lead to bacterial hemorrhagic septicemia (BHS),6 exercise Aeromonas septicemia (MAS)7 and epidemic ulcer syndrome (EUS).8 Among these infections, Aeromonas veronii (A. veronii), identified as an opportunistic pathogen,9,10 has demonstrated its ability to infect a diverse range of fish species, such as cyprinid fish,11 Largemouth bass (Micropterus salmoides),12 freshwater dark sleeper (Odontobutis potamophila),13 Nile tilapia (Oreochromis niloticus),14 and crucian carp (Carassius carassius),15 among others. Recent research has indicated a potential association between A. veronii and alterations in intestinal flora.16
The gut microbiota comprises various microorganisms, including probiotics, pathogens, and neutral bacteria.17,18 These microorganisms play important roles in regulating the host’s physiology, immunity, and nutrition.19–21 Previous studies have demonstrated a strong connection between alterations in intestinal flora and numerous diseases, including diarrhea, obesity, diabetes, and cancer.22–25 Several studies have also revealed that pathogen infections can alter the composition of fish intestinal microflora.26–28 The intestinal tract may also serve as a potential source or route for pathogenic infections.29,30 Simultaneously, Recent research has highlighted the significant role of probiotics as alternatives to antibiotics for disease control, which have shown substantial effects in modulating gut microbiota, promoting host growth and metabolism, and inhibiting pathogens.31–33 Lactobacillus plantarum (L. plantarum), a Gram-positive lactic acid bacterium, has demonstrated its potential as a probiotic, particularly in suppressing pathogenic bacteria and regulating host gut microbiota.34 Studies have shown that including L. plantarum in feed can enhance the growth of Nile tilapia and increase resistance to Aeromonas hydrophila infection.35 In the large yellow croaker (Larimichthys crocea), L. plantarum has exhibited significant antimicrobial activity against several aquaculture-related pathogens.36 Moreover, bacteriocins produced by L. plantarum effectively inhibit the growth of Staphylococcus aureus, Listeria monocytogenes and other bacteria.37 These findings demonstrate the potential of L. plantarum as a probiotic in aquaculture, offering a viable alternative to antibiotics for disease prevention and control.
Lateolabrax maculatus (L. maculatus) is widely distributed in East Asia, extending to the border between China and Vietnam in the south to the southeast coast of South Korea in the north.38 This species holds significant economic importance and has gained prominence as one of the most promising breeds owing to the rapid growth of the L. maculatus culture industry.39 However, L. maculatus is susceptible to Aeromonas disease caused by A. veronii.40 Currently, most research on fish disease resistance mechanisms mainly focuses on physiological and transcriptional aspects post-infection, with limited exploration of the role of intestinal microorganisms in disease resistance. There have been limited studies on the composition and structure of the intestinal microflora of L. maculatus during bacterial infection. Additionally, the impact of L. plantarum on the gut microbiota of L. maculatus remains unexplored. Therefore, this study aimed to investigate the composition and differences in the intestinal flora of healthy and A. veronii-infected L. maculatus, evaluate the microbiota changes caused by the disease, and assess the effect of L. plantarum on the gut microbiota of L. maculatus during bacterial infection. Based on the previous information, we harvested intestinal tissue samples from healthy L. maculatus at various time points after injection of A. veronii, preceded by the pretreatment with L. plantarum. This study holds the potential to provide theoretical insights into enhancing the prevention and control of bacterial infections and the development of probiotics within large-scale, high-density, and intensive cultures of L. maculatus.
Materials and Methods
Ethics statement
L. maculatus is neither an endangered nor a protected species, and conducting experiments with this species in China does not require permission. The animal experiment was approved by the Animal Ethics Committee of the Chinese Academy of Fishery Sciences (Approval No. 2011AA1004020012).
Experimental animals, bacterial, and sample collection
Healthy L. maculatus specimens were sourced from the Zhuhai Experimental Base of the South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (Guangzhou, China). Prior to infection, L. maculatus (65 ± 3 g) were randomly distributed into two separate culture buckets (500 L) with 30 fish in each bucket. They were maintained in aerated freshwater at 29 ± 1 °C, with one-third of the water being replaced daily.
Feeding was stopped 24 h before the start of the experiment. In the bacterial challenge experiment, the entire intestinal tissues of three fish were initially extracted from each of the two culture buckets. Subsequently, the remaining fish in both culture buckets were injected intraperitoneally with A. veronii at a dose of 8.5 × 108 CFU/g. The entire intestinal tissues were collected from L. maculatus at two distinct time points (24 and 48 h) post-injection (three biological replicates at each time point). All collected samples were frozen in liquid nitrogen and stored at -80 °C for further analysis. The A. veronii colonies utilized in the challenge tests were preserved in our laboratory. The L. plantarum used in the experiment is a commercially available strain commonly used as a dietary supplement.
DNA extraction
Microbial DNA was extracted using the HiPure Soil DNA Kit or HiPure Stool DNA Kit (Magen, Guangzhou, China) according to the instructions of the manufacturer. The V3–V4 region of the 16S rDNA target, part of the ribosomal RNA gene, was amplified by polymerase chain reaction (PCR) (95 °C for 5 min, followed by 30 cycles at 95 °C for 1 min, 60 °C for 1 min, and 72 °C for 1 min and a final extension at 72 °C for 7 min) using primers 341F (CCTACGG GNGGCWGCAG) and 806R (GGACTACHVGGGTATCTAAT). The PCR reaction mixture consisted of 50 μL, containing 10 μL of 5 × Q5@ Reaction Buffer, 10 μL of 5 × Q5@ High GC Enhancer, 1.5 μL of 2.5 mM deoxynucleotide triphosphates (dNTPs), 1.5 μL of each primer (10 μM), 0.2 μL of Q5@ High-Fidelity DNA Polymerase, and 50 ng of template DNA. All PCR reagents were purchased from New England Biolabs (USA). Amplicons were extracted from 2% agarose gels, purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, USA) according to the instruction of the manufacturer, and quantified using the ABI StepOnePlus Real-Time PCR System (Life Technologies, USA). The purified amplicons were combined in equimolar proportions and subjected to paired-end sequenced (PE250) on an Illumina 2500 platform according to standard protocols.
Bioinformatics analysis
After sequencing, raw reads were filtered using FASTP41 (version 0.18.0) to obtain high-quality clean reads. Subsequently, clean paired-end reads were merged into raw tags using FLASH42 (version 1.2.11) designed for accurate merging based on overlap analysis. Noisy sequences were removed to obtain high-quality clean tags using specific filtering conditions.43 Utilizing the UPARSE pipeline44 (version 9.2.64), the clean tags were clustered into operational taxonomic units (OTUs) with a similarity threshold of ≥ 97%. All chimeric tags were removed using the UCHIME algorithm.45 Representative sequences for each OTU were selected based on the highest abundance tag sequence. The representative OTU sequences were classified using a naive Bayesian model in the RDP classifier46 (version 2.2) based on the SILVA database47 (version 132), with a confidence threshold of 0.8. Abundance statistics for each taxonomy were visualized using Krona48 (version 2.6). Community composition was visualized using stacked bar plots in R with the ggplot2 package49 (version 2.2.1), and species abundance heatmaps were created using the pheatmap package50 (version 1.0.12) in the R project. Venn diagrams for group comparisons were generated using the VennDiagram package51 (version 1.6.16) in R. Alpha diversity metrics, including Chao1, Shannon, Simpson, and Good’s coverage, were computed to assess species richness and evenness within microbial communities using the vegan package in QIIME52 (version 1.9.1). including Adonis (PERMANOVA) and Anosim tests, were performed using the vegan package in R53 (version 2.5.3) and plotted in R project ggplot2 package49 (version 2.2.1). Statistical analysis of Adonis (also called Permanova) and Anosim test was calculated in R project Vegan package53 (version 2.5.3). Diversity indices and bacterial abundances among different groups were compared using one-way ANOVA. Statistical significance was set at P < 0.05.
Results
Sequences analyses
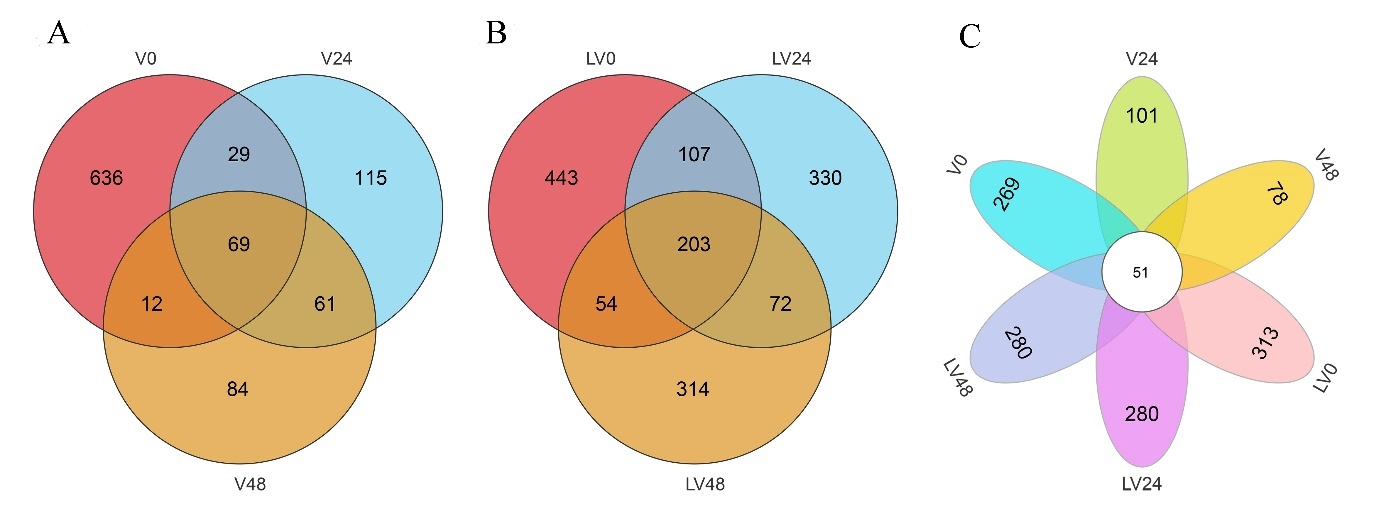
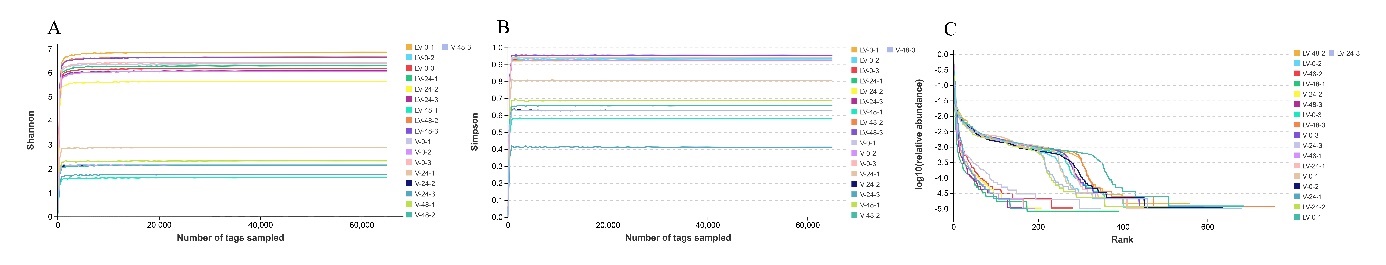
High-throughput sequencing was used to analyze sample sequences. After data optimization, 303,793 effective sequences were obtained from the 18 intestinal samples of L. maculate (Table. 1). Based on 97% sequence identity, the effective sequences were divided into 2,845 OUTs. The number of OTUs per sample varied, ranging from 161 to 760 (Table 1). Moreover, in the infection groups (V0, V24, and V48), 69 common OTUs were identified, while in the infection groups after pretreatment with L. plantarum (LV0, LV24, and LV48), 203 common OTUs were observed (Fig. 1 A and B). There were 51 common OTUs among the six different small groups (Fig. 1C). The saturation observed in the Shannon, Simpson, and rank abundance curves suggests that the sequencing depth, abundance, and homogeneity adequately capture gut microbiota diversity in the samples (Fig. 2A–C).
.jpeg)

Analysis of the diversity of the intestinal microbial community
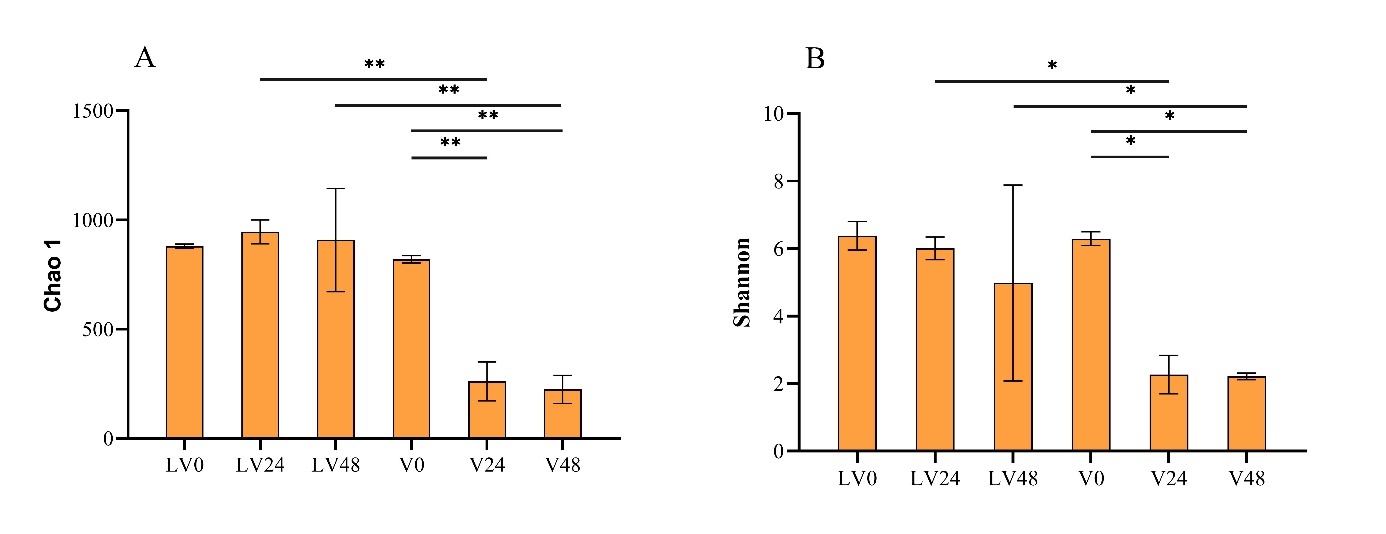
To assess the alpha diversity of gut microbial communities in various treatment groups and time points, Chao1, Shannon, and Good’s coverage indices were used. The Good’s coverage estimates across all samples ranged from 99.74% to 99.96%, indicating exceptional coverage (Table 2). Figure 3A presents the mean value of the Chao1 index, which was significantly higher in the V0 group than in the V24 (p < 0.01) and V48 groups (p < 0.01), indicating a significant difference in intestinal flora abundance between the V0 group and the other two groups. Similarly, the mean Chao 1 index was significantly higher in the LV24 group than in the V24 group (p < 0.01), indicating a significant difference in gut microbe abundance between these two groups. Furthermore, the mean Chao1 index was significantly higher in the LV48 group than in the V48 group (p < 0.01). Additionally, the mean Shannon’s index was significantly higher in the V0 group than in the V24 and V48 groups (p < 0.05). This indicated a significant difference in the homogeneity of the gut microbes between the V0 group and the other two groups. The Shannon index was significantly lower in the V24 group than in the LV24 group (p < 0.05). A similar significant difference was observed in the V48 group compared with the LV48 group (p < 0.05) (Fig. 3B). However, the LV24 and LV48 groups showed large dispersion, and PcoA indicated a significant difference in the gut microbiota composition across all six groups (Adonis, R2 = 0.569, p < 0.001), with a 53.39% distance in PCo1. Notably, the LV24 and LV48 groups formed a cluster in PCo1, the V24 and V48 groups clustered together in PCo1, and the V0 group clustered with the LV0 group (Fig. 4A–B).


Composition analysis of the gut microbiota community structure at the phylum level in different groups
The gut microbiota community structures of the different treatments were evaluated at the phylum level based on the sampling time. Among the 27 bacterial phyla identified, with a few unclassified OTUs, the dominant phyla in the gut microbiota of the V0 group (healthy fish) were Firmicutes (58.98%), Bacteroidetes (18.35%), Proteobacteria (9.92%), and Actinobacteria (5.72%) (Fig. 5A). The relative abundance of Proteobacteria increased significantly at 24 h (p < 0.01), whereas the relative abundances of Firmicutes (p < 0.001) and Bacteroidetes (p < 0.001) decreased significantly (Fig. 6A–C). In the L. plantarum pretreatment group, the dominant phyla in the gut microbiota of the LV0 group (healthy fish) were Firmicutes (55.61%), Bacteroidetes (19.84%), Proteobacteria (12.31%), and Actinobacteria (6.43%) (Fig. 5A). These were similar to the composition of the V0 group; however, their relative abundances differed. The relative abundances of Proteobacteria, Firmicutes, and Bacteroidetes remained stable over time; however, a notable contrast was observed in the untreated diseased fish. Specifically, the untreated diseased fish displayed a higher relative abundance of Proteobacteria, while the relative abundances of Firmicutes (p < 0.001) and Bacteroidetes (p < 0.01) were significantly lower compared with those in the diseased fish in L. plantarum pretreatment group (Fig. 6A–C).


The top 20 bacterial genera in each group are shown in the abundance heatmap (Fig. 5B). The results from Fig. 5B and C indicate that the dominant flora in both healthy fish and those treated with L. plantarum were the same, although the relative abundances differed. Over time, apart from a few bacterial genera (mostly pathogenic bacteria), the relative abundance of bacterial genera in the untreated diseased fish significantly decreased than that in healthy fish. Most of the bacterial genera in the untreated diseased fish exhibited a downward trend; however, the overall downward trend was lower than that in the diseased fish after L. plantarum treatment. Among them, the relative abundance of Aeromonas significantly increased at 24 h (p < 0.001) and decreased at 48 h (p < 0.01) in both untreated infected fish and L. plantarum-treated fish. The relative abundance of Aeromonas was significantly lower in the LV48 group than in the V48 group (p < 0.001) (Fig. 6D). For Vibrio, the relative abundance increased significantly over time in untreated infected fish (p < 0.05) compared with healthy fish, however, there was no significant change in infected fish treated with L. plantarum (Fig. 6E). Regarding Acinetobacter, the relative abundance in untreated diseased fish increased significantly increased at 24 h (p < 0.001) and decreased at 48 h (p < 0.001), while that of Acinetobacter in diseased fish treated with L. plantarum showed no significant change (Fig. 6F). The relative abundance of Lactobacillus and Bifidobacterium in untreated diseased fish significantly decreased over time than that in healthy fish (p < 0.05), but there was no significant difference in the relative abundance of Lactobacillus and Bifidobacterium in L. plantarum-treated diseased fish (Fig. 6G and H). The relative abundances of Vibrio, Acinetobacter, Lactobacillus, and Bifidobacterium in untreated diseased fish were significantly different from those in L. plantarum-treated diseased fish (p < 0.05) (Fig. 6E–H).
Discussion
Aeromonas spp. have consistently posed significant threats in aquaculture, causing infectious diseases that result in significant economic losses globally.54 In recent years, the role of the gut microbiota in host disease resistance has become a frontier of research. Many studies have demonstrated the connection between various diseases and alterations in the intestinal microflora.32,55,56 The gut microbiota plays a crucial role in preventing and treating diseases by regulating the intestinal barrier and environment, thereby reducing the invasion and colonization of pathogens. Given the growing threat posed by infectious pathogen invasion in the fish culture industry, it is essential to explore the relationship between pathogen invasion and the diversity of the intestinal flora. Such exploration is vital for understanding how intestinal microorganisms affect host health and for developing effective measures to prevent or treat diseases. Moreover, probiotics have been demonstrated to stimulate host growth and development, boost host immunity, and potentially replace antibiotics.57 Using high-throughput sequencing, this study compared and analyzed the diversity and dynamic changes in the intestinal flora of L. maculatus infected with A. veronii and L. maculatus infected with A. veronii after pretreatment with L. plantarum. This study aimed to investigate the effects of A. veronii infection on the intestinal flora of L. maculatus and the modulation of the intestinal flora of L. maculatus by L. plantarum.
The Chao 1 index, Shannon index, and PCoA analyses revealed distinct patterns in this study. The untreated infected group exhibited a significant decrease in diversity over time. In contrast, the diversity of the infected group with L. plantarum pretreatment remained relatively stable over time, and the difference in diversity between the two groups was significant. These findings suggest that A. veronii infection reduces the abundance and evenness of microbial diversity in L. maculatus, while L. plantarum contributes to the preservation of microbial diversity. Previous research has indicated that a higher diversity of gut microbes is advantageous for maintaining dynamic intestinal balance and function.58
Maintaining the “microbial balance” in the intestinal tract is vital to health. When analyzing the gut microbiota composition in L. maculatus, Firmicutes, Bacteroidetes, and Proteobacteria emerged as the dominant phyla, aligning with previous findings on fish intestinal flora.59 Proteobacteria constituted the largest phylum in infected fish, encompassing well-studied pathogens and indicating dysbiosis in the gut microbiota.60,61 The increase in the relative abundance of Proteobacteria may contribute to the development of inflammatory bowel disease.62 These findings show that a higher Proteobacteria abundance heightens the susceptibility of fish to pathogenic infections. Additionally, Bacteroidetes was also a dominant phylum in the intestinal flora of healthy fish. Studies have shown that Bacteroides is related to the intestinal immune response,63,64 and a higher abundance of Bacteroidetes helps to improve host immunity and contributes to the dynamic balance of gut microbiota.65 In our study, compared with healthy fish, infected fish exhibited an increase in Proteobacteria abundance and a decrease in Bacteroidetes abundance over time. This suggests an imbalance in the intestinal flora of A. veronii-infected L. maculatus. In contrast, diseased fish treated with L. plantarum showed a lower abundance of Proteobacteria and a higher abundance of Bacteroidetes than those in untreated diseased fish. This indicates that L. plantarum treatment is beneficial in reducing infection risk and maintaining intestinal flora balance.
At the genus level, A. veronii infection increased the abundance of some pathogenic bacteria and reduced the abundance of many beneficial bacteria compared to healthy fish. This disturbance in intestinal flora’s environmental balance may reduce the intestinal tract’s resistance to pathogenic bacteria. In contrast, fish pretreated with L. plantarum exhibited a more stable intestinal flora, highlighting the role of L. plantarum in preserving microbial diversity during prevention and antibacterial treatment. Comparatively, untreated diseased fish displayed higher Aeromonas, Vibrio, and Acinetobacter levels and lower Lactobacillus and Bifidobacterium levels. While the Aeromonas content was higher in diseased fish with L. plantarum pretreatment than in healthy fish, there were no significant changes in Vibrio, Acinetobacter, Lactobacillus, and Bifidobacterium. Additionally, the Vibrio, Acinetobacter, Lactobacillus, and Bifidobacterium levels in untreated diseased fish were lower than those in diseased fish with L. plantarum pretreatment. Changes in Aeromonas may directly result from A. veronii infection, as indicated by a similar study.66 Additionally, Vibrio, an opportunistic pathogen prevalent in freshwater and seawater, is a leading cause of Vibrio disease in aquaculture,67 presenting symptoms such as fish septicemia and gastroenteritis, among other symptoms.68 Acinetobacter, identified as a fish pathogen, poses a new threat to the aquaculture industry, leading to severe septicemia outbreaks in fish farms.69 An imbalance in the intestinal microbial structure may have caused the increase in relative abundance of Vibrio and Acinetobacter. Lactobacillus and Bifidobacterium are probiotic bacteria. Lactobacillus promotes host growth and reproduction, improve immunity, and disease resistance,70–72 while Bifidobacterium induces better growth of trout seedlings and increase digestion and nutritional utilization.73 In addition, CpG oligodeoxynucleotides (ODNs) from Bifidobacterium may serve as immunostimulatory modulators of the immune response.74 LAB combinations (Lactobacillus and Bifidobacterium) can improve the host’s growth performance and intestinal health and reduce the risk of pathogenic infections.75 Therefore, it is speculated that A. veronii infects the intestine and suppresses some beneficial bacteria, such as Lactobacillus and Bifidobacterium. This suppression could create an environment where Aeromonas and other pathogenic bacteria occupy more niches, ultimately disrupting the balance of intestinal flora. Treatment with L. plantarum helps maintain the dynamic balance of intestinal flora and potentially protects sea bass.
In conclusion, this study investigates the composition and differences in the gut microbiota of healthy and A. veronii-infected L. maculatus to assess the microbial alterations caused by the infection and evaluate the influence of L. plantarum during the bacterial infection process. The findings indicate that A. veronii infection disrupts the gut microbiota equilibrium in L. maculatus. L. plantarum may stabilize the gut microbiota, thereby providing protection against A. veronii infection. These results offer theoretical support for developing strategies to prevent bacterial infections and promote the use of probiotics in the intensive, high-density aquaculture of L. maculatus.
Acknowledgments
This work was supported by the Central Public-interest Scientific Institution Basal Research Fund, CAFS(NO. 2024XT02), the Central Public-interest Scientific Institution Basal Research Fund, CAFS (No. 2023TD21), the National Key Research and Development Program of China (No. 2022YFD2400503), Guangdong Province Strategic Projects for Rural Revitalization (No. 2022-spy-00-009), and the Guangdong Basic and Applied Basic Research Foundation (No. 2023A1515030022).
Authors’ Contribution – CRediT taxonomy
Methodology: Changhong Lin (Lead). Formal Analysis: Changhong Lin (Equal), Lihua Qiu (Equal), Bo Zhang (Equal). Investigation: Changhong Lin (Equal), Lihua Qiu (Equal), Bo Zhang (Equal). Writing – original draft: Changhong Lin (Lead). Resources: Pengfei Wang (Equal), Bo Zhang (Equal). Writing – review & editing: Lulu Yan (Equal). Conceptualization: Chao Zhao (Lead). Funding acquisition: Chao Zhao (Lead). Supervision: Chao Zhao (Lead).
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
Research involving animals
The animal experiment was approved by the Animal Ethics Committee of the Chinese Academy of Fishery Sciences (Approval No. 2011AA1004020012).
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
The data that has been used is confidential.