Introduction
The Chinese pearl oyster (Pinctada fucata) belongs to the phylum Mollusca, class Lamellibranchia, subclass Pteriomorphia, order Pterioida, and family Pteriidae.1 It holds a pivotal position in the pearl cultivation industry and is widely distributed in the tropical and subtropical waters of Guangdong, Guangxi, and Hainan in China.2 As one of the primary species cultivated for seawater pearls in China, Chinese pearl oysters contribute to over 95% of the country’s total marine pearl output.3 Current research on Chinese pearl oysters predominantly concentrates on seedling breeding, genetic breeding, and molecular markers, with comparatively less attention directed towards reproductive characteristics, breeding strategies, and germplasm conservation.4 In the artificial breeding process of Chinese pearl oysters, common methods like air exposure and chemical induction are employed for sperm and egg collection.5 However, the inability to discharge sperm normally or the asynchrony in the release of sperm and eggs often occur, it becomes imperative for the sperm to remain viable and maintain a certain level of activity for successful fertilization.6
Exogenous additives are often utilized to influence the activation rate and active duration of sperm.7 Trehalose, as a non-reducing sugar, can create a unique protective film within cells to prevent damage caused by dehydration, thereby maintaining cell structural integrity and providing antioxidant protection.8 It has been reported that adding 60 mmol·L-1 of trehalose to the sperm samples of Amur sturgeon (Acipenser schrenckii) can protect the sperm to a certain extent and significantly enhance its vitality.9 Bovine serum albumin, serving as a natural antioxidant and nutrient carrier, can supply essential nutrients while reducing free radical damage and protecting sperm from oxidative stress.10 Studies have shown that using a 20% bovine serum diluent for preserving the sperm of Hong Kong oyster (Magallana hongkongensis) can significantly prolong sperm lifespan.11 Fructose and glucose, as primary nutrients within the sperm’s protoplasm and as exogenous energy sources, are crucial for powering sperm vitality. The insufficient levels of fructose and glucose in seminal plasma can directly reduce sperm motility capabilities.12 The primary energy metabolism pathway for sperm is glycolysis, the process by which sperm, under aerobic or anaerobic conditions, breaks down seminal plasma sugars such as fructose, glucose, and mannose to produce pyruvate or lactate. Research has shown that fructose serves as a major energy source for sperm, which also contains a small amount of glucose.9 These monosaccharides not only regulate osmotic pressure but can also penetrate sperm cells to serve as energy substrates, producing ATP through glycolysis or the tricarboxylic acid cycle to fuel sperm activity.13 Studies have confirmed that adding fructose to the diluent protects Japanese pearl oyster sperm.14
ATP hydrolysis rates and energy metabolism of sperm may be influenced by temperature and ion concentration, affecting its vitality and movement. Elevated temperatures increase metabolic levels and ATP hydrolysis rates, intensifying sperm movement. Since sperm cells have limited energy reserves, excessive temperatures can irreversibly inhibit sperm activity or even cause premature death. Conversely, lower temperatures may reduce sperm motility and intensity but slow down ATP hydrolysis, extending sperm lifespan.15 Studies have shown that sperm of longfin dace (Rhinogobio ventralis) stored at 4°C exhibit longer movement phases and lifespans compared to those stored at 20°C.16,17 It has been reported that storing silver bream (Rhabdosargus sarba) sperm between 3 to 5°C extends its lifespan compared to room temperature conditions.18 Additionally, adding ammonia can provide a more suitable environment for glycolysis.19 Studies have shown that ammonia seawater is commonly used in breeding and genetic research in aquatic animals.20 According to reports, adding natural seawater to 150 μL of 25% concentrated ammonia water to dilute it to 1 L and thoroughly mixing, method is utilized for the activation of giant pearl oyster sperm, effectively extending the sperm survival time by 4 to 8 hours.21 In activation experiments conducted on glass slides, the movement of sperm ceases when NH4+ is converted into ammonia and evaporates. When NH4+ is reintroduced to the environment, the sperm resumes movement. This observation underscores the critical role of NH4+ in sperm activation, as NH4+ is capable of maintaining the ion channels in a continuous open state. The ability of NH4+ to keep these channels open facilitates the essential physiological processes for sperm motility, indicating that the presence and maintenance of ammonium ions are vital for the active state and functionality of sperm.
This research focuses on the fresh sperm of Chinese pearl oyster, exploring the impact of exogenous additives under different temperature conditions and with or without the addition of ammonia water on sperm vitality in the short term. The goal is to provide a theoretical basis for the subsequent ultra-low temperature long-term preservation of Chinese pearl oyster, aiming to address the practical issues currently faced in the artificial breeding process of Chinese pearl oyster and offer scientific references for germplasm conservation, artificial fertilization, and seedling cultivation of Chinese pearl oyster.
Materials and Methods
Experimental Animals
The Chinese pearl oysters were sourced from the Tropical Aquaculture Research and Development Center of the South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, situated in Xincun Port, Lingshui Li Autonomous County, Hainan Province, China. The one-year-old Chinese pearl oysters were chosen for this study. Throughout the sperm collection process, the oysters were temporarily housed in seawater with a salinity of 35±0.2 and a temperature of 25±2°C for standby use.
Trait Data Collection
Seventy Chinese pearl oysters were selected, and among them, three male individuals with the most developed gonads and highest maturity were chosen. Measurements and weighing were conducted for shell length, shell height, shell width, wet weight, soft body weight, and testis weight. The results are as follows (Table 1).
Sperm Collection
From a pool of 70 Chinese pearl oysters, 3 male individuals exhibiting the fullest and most mature gonads were carefully selected. Utilizing a sterile scalpel, one side of the adductor muscle of each oyster was incised to expose the visceral mass, with the gonads nearly encompassing the entire visceral sac and presenting a white or pale-yellow appearance. Subsequently, the gonads were meticulously dissected, and a clean absorbent paper was employed to thoroughly absorb the mixture of seawater and tissue fluid.
The obtained Chinese pearl oyster gonads were then positioned on a 400-mesh sieve cloth, where they were finely minced with scissors and rinsed with natural seawater to eliminate excess tissue fragments. This process resulted in a relatively clean sperm suspension, which was subsequently diluted twice. The suspension was carefully transferred into a clean beaker for future use.
Additives on the Vitality of Chinese pearl oyster Sperm
Natural seawater served as the control group, while the experimental groups were categorized into four treatment groups. The seawater used in the experiment was collected from Xincun Port, Lingshui Li Autonomous County, Hainan, China. The salinity of the seawater was (35±0.2)‰, and the temperature was (25±2)°C. The concentrations of additives used in this experiment were selected after several preliminary experiments. It was found that different additives yielded the best results under the condition of 2.8 mmol/L. In the experimental groups, 2.8 mmol/L of trehalose,14 bovine serum albumin,22 fructose,12 and glucose23 (The D-(+)-trehalose, bovine serum albumin (BSA), D-fructose, and D-glucose used in the experiment were all products of Shantou Xilong Chemical Co., Ltd. in China and were domestically produced analytical grade reagents.) (Table 2) were dissolved in natural seawater, each added separately. The semen and the dissolved additives were then diluted at a 1:20 (V:V) ratio to create experimental group samples. In the control group, semen was diluted with seawater without additives at a 1:20 (V:V) ratio. After thorough mixing of the diluted samples, they were dispensed into 15 mL plastic centrifuge tubes, with 10 mL per tube. At 0, 2, 4, 8, 12, 16, 20, and 24 hours—comprising a total of 8-time points. A 2 μL samples were extracted from each time point, smeared into a sperm counting chamber, and sperm vitality was observed and recorded using a sperm quality analyzer (Beionmed S3 model, produced by Shanghai Beion Medical Ltd., China). Each group was replicated three times in parallel.
Temperature on the Vitality of Chinese pearl oyster Sperm
Expanding on the selection of optimal additives, each experimental group underwent treatment under different room temperature (25°C) and low temperature (4°C), resulting in the division of the experimental setup into two distinct groups (Table 3). In the experimental groups, 2.8 mmol/L of fructose and glucose, dissolved in natural seawater, were separately added. The semen and the dissolved additives were then diluted at a 1:20 (V:V) ratio to produce experimental group samples. These samples were dispensed into 15 mL plastic centrifuge tubes, with 10 mL per tube, and were subjected to either room temperature or low-temperature conditions. At 0, 2, 4, 8, 12, 16, 20, and 24 hours—amounting to a total of 8-time points—2 μL samples were obtained from each time point, smeared into a sperm counting chamber, and sperm vitality was observed and recorded using a sperm quality analyzer. Each group was replicated three times in parallel.
Ammonia Addition on the Vitality of Chinese pearl oyster Sperm
Building upon the outcomes of the temperature experiment, under low-temperature conditions (4°C), the experimental groups were subdivided into two categories (refer to Table 4). In the experimental groups, 2.8 mmol/L of fructose and glucose, dissolved in natural seawater, were individually added. The semen and the dissolved additives were then diluted at a 1:20 (V:V) ratio. Concurrently, 2 mmol/L of NH4+ seawater, prepared by diluting 150 μL of 25% concentrated ammonia solution with natural seawater to 1L, thoroughly mixed, freshly prepared, and used for activating the Chinese pearl oyster sperm. The 25% concentrated ammonia solution used in the experiment is a product of Shantou Xilong Chemical Co., Ltd. in China and is a domestically produced analytical grade reagent, was introduced. This combined mixture served as the experimental group sample. The samples were dispensed into 15 mL plastic centrifuge tubes, with 10 mL per tube, and were subjected to either room temperature or low-temperature conditions. At 0, 2, 4, 8, 12, 16, 20, and 24 hours—comprising a total of 8 time points—2 μL samples were procured from each time point, smeared into a sperm counting chamber, and sperm vitality was observed and recorded using a sperm quality analyzer. Each group was replicated three times in parallel.
Sperm Vitality Assessment and Quality Evaluation
The experiment design encompasses eight observation and recording time points: 0, 2, 4, 8, 12, 16, 20, and 24 hours. The analysis of Chinese pearl oyster sperm vitality is conducted utilizing a Computer-Assisted Sperm Analysis (CASA) system (Ming Hao Technology Co., Ltd., Huizhou, China). To ensure consistency, the number of captured sperm in each lens view is maintained between 400 to 600. For each sample, six random fields of view are selected for sperm vitality testing. Sperm vitality, denoted as sperm motility (Progressive Motility + Non-Progressive Motility, %).
Data Analysis
The data of sperm parameters measured by the Computer-Assisted Sperm Analysis (CASA) system during the experiment were statistically analyzed using SPSS 27.0. One-way Analysis of Variance (ANOVA) was conducted with the software, where p<0.05 indicated a significant difference, p<0.01 indicated a highly significant difference and p<0.001 indicated an extremely significant difference. Results were presented as mean±standard deviation (mean±S.D). Graphs were created using Origin 2021.
Results
The Impact of Additives on the Vitality of Chinese pearl oyster Sperm
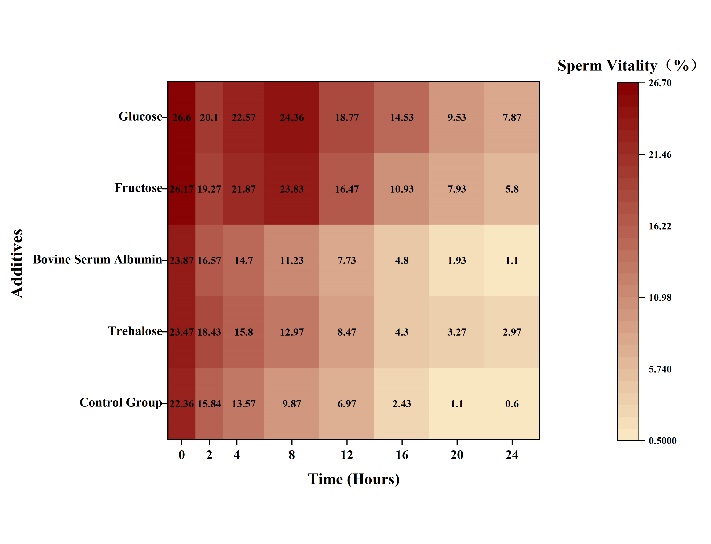
After sperm activation (Figure 1), the sperm motility of all experimental groups was similar to that of the control group at 0 hours, with no significant difference (p>0.05). Among them, glucose showed a decreasing trend in sperm motility 2 hours after activation. From 2 to 8 hours after activation, sperm motility showed an increasing trend, and reached a peak value of 24.36% at 8 hours after activation, showing a significant difference compared to the control group’s sperm motility (p<0.05). The mean decrease in sperm viability was the greatest at 8 to 12 hours after activation, which was 5.59%, with a significant decreasing trend (p<0.05) compared to other periods. From 12 to 24 hours after activation, sperm motility showed a gradually declining trend. Moreover, from 4 to 24 hours after activation, compared to the control group, the impact on sperm motility showed a highly significant difference (p<0.01).

Fructose showed a decreasing trend in sperm motility 2 hours after activation. From 2 to 8 hours after activation, sperm motility showed an increasing trend, and reached a peak value of 23.83% at 8 hours after activation, showing a significant difference compared to the control group’s sperm motility (p<0.05). The mean decrease in sperm viability was the greatest at 8 to 12 hours after activation, which was 7.37%, with a significant decreasing trend (p<0.05) compared to other periods. From 12 to 24 hours after activation, sperm motility showed a gradually declining trend. Moreover, from 4 to 24 hours after activation, compared to the control group, the impact on sperm motility showed a highly significant difference (p<0.01).
The control group, trehalose, and bovine serum albumin experimental groups showed a gradually declining trend from 0 to 24 hours after activation, and the greatest decrease in sperm motility occurred at 2 hours. Trehalose and bovine serum albumin showed no significant difference in sperm motility at any time point compared to the control group (p>0.05).
The Impact of Temperature on the Vitality of Chinese pearl oyster Sperm
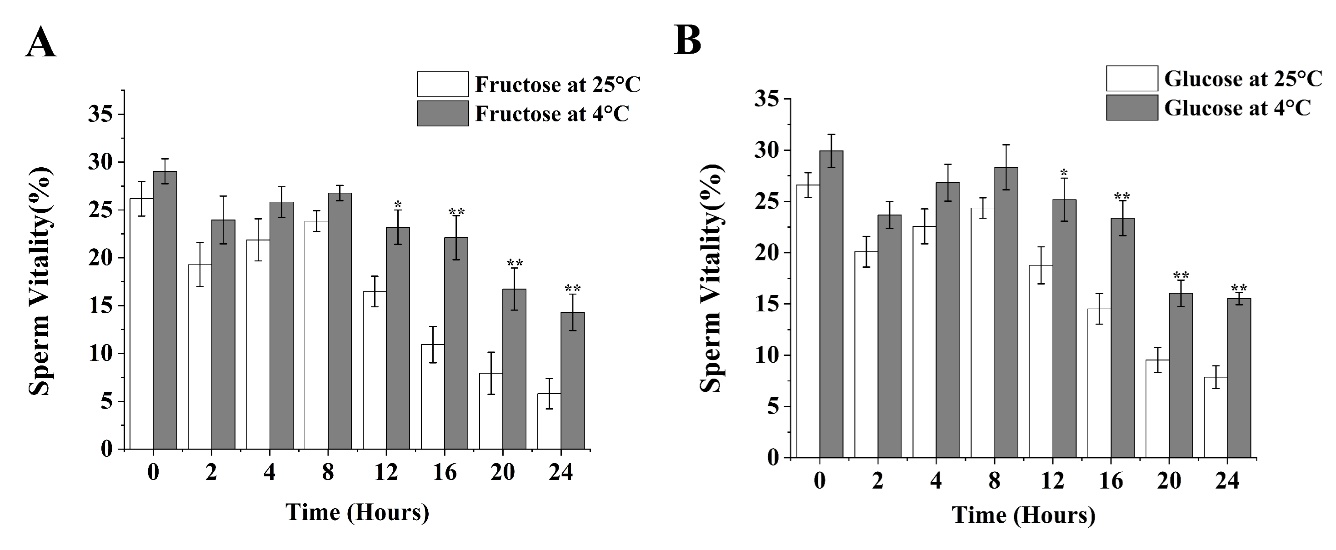
Under 4°C conditions, fructose (Figure 2A) saw sperm motility reaching its first peak value of 29.04% at 0 hours after activation. Sperm motility then showed a decreasing trend after 2 hours of activation. From 2 to 8 hours after activation, sperm motility showed an increasing trend and reached its second peak after 8 hours. From 8 to 24 hours after activation, sperm motility showed a gradually decreasing trend, with motility reaching its lowest value of 14.3% at 24 hours after activation. Notably, at 12 hours after activation, sperm motility under 4°C conditions showed a significant difference compared to 25°C conditions (p<0.05), and from16 and 24 hours after activation, the difference in sperm motility between 4°C and 25°C conditions was highly significant (p<0.01). Specifically, the average rate of decrease of 5.33% in sperm motility was the greatest between 16 to 20 hours after activation under 4°C conditions, showing a significant declining trend compared to other adjacent time periods (p<0.05).

For glucose (Figure 2B) under 4°C conditions, sperm motility reached its first peak value of 29.93% at 0 hours after activation. After 2 hours of activation, sperm motility showed a decreasing trend. From 2 to 8 hours after activation, sperm motility displayed an increasing trend and reached its second peak after 8 hours. From 8 to 24 hours after activation, sperm motility consistently showed a gradually decreasing trend, with motility reaching its lowest value of 15.53% at 24 hours after activation. Notably, at 12 hours after activation, sperm motility under 4°C conditions showed a significant difference compared to 25°C conditions (p<0.05), and from 16 and 24 hours after activation, the difference in sperm motility between 4°C and 25°C conditions were highly significant (p<0.01). Specifically, the average rate of decrease of 7.34% in sperm motility was the greatest between 16 to 20 hours after activation under 4°C conditions, showing a significant declining trend compared to other adjacent time periods (p<0.05).
The Impact of Ammonia Addition on the Vitality of Chinese pearl oyster Sperm
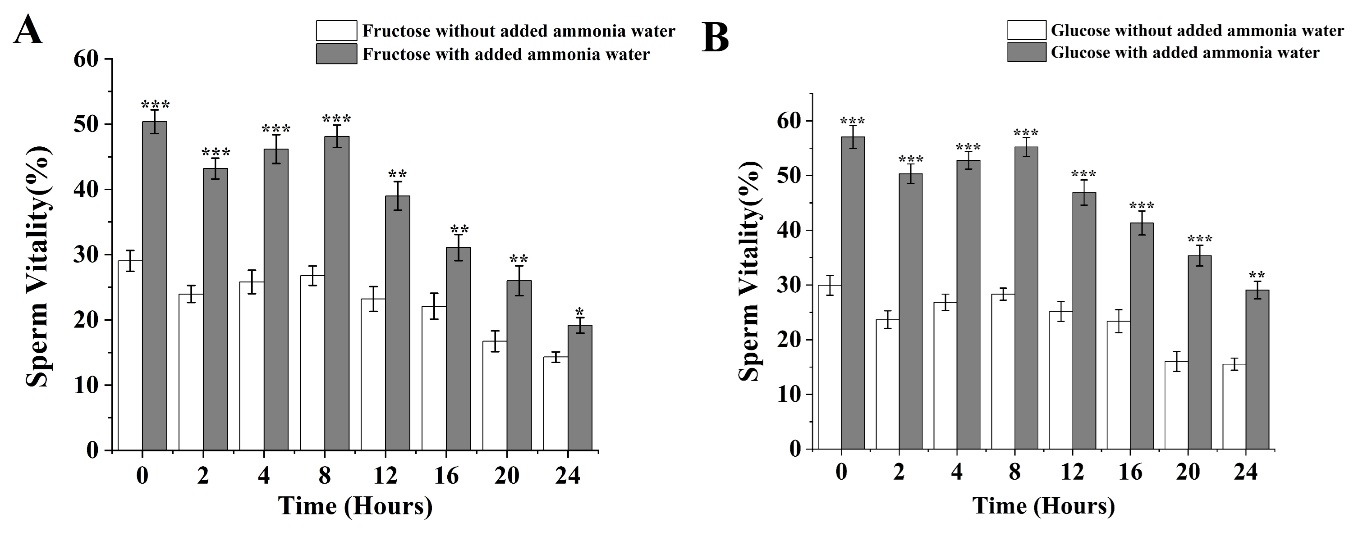
Analyzing the impact of adding ammonia to fructose and glucose on sperm motility under low temperature (4°C) conditions over a 24-hour period reveals distinct effects. For fructose (Figure 3A) under 4°C conditions with added ammonia, sperm motility reached a peak value of 50.37% immediately at 0 hours post-activation. After 2 hours, a downward trend in sperm motility was observed, followed by an upward trend from 2 to 8 hours. Sperm motility gradually declined from 8 to 24 hours, reaching the lowest point of 19.17% at 24 hours. Notably, from 0 to 8 hours post-activation, the difference in sperm motility compared to samples without added ammonia at the same temperature was highly significant (p<0.001). Between 12 to 20 hours, a highly significant difference was also observed compared to samples without ammonia (p<0.01), and at 24 hours, the difference remained significant (p<0.05). The greatest average decrease of 6.73% in sperm motility was seen from 8 to 12 hours, showing a significant decline compared to adjacent time periods (p<0.05).

For glucose (Figure 3B) under 4°C conditions with added ammonia, sperm motility reached a peak 57.07% at 0 hours post-activation. After 2 hours, sperm motility showed a decreasing trend, followed by an increasing trend from 2 to 8 hours. Sperm motility gradually declined from 8 to 24 hours, reaching the lowest point of 19.17% at 24 hours. Between 0 to 20 hours post-activation, the difference in sperm motility compared to samples without added ammonia at the same temperature was highly significant (p<0.001). At 24 hours, a highly significant difference was noted compared to samples without ammonia (p<0.01). The greatest average decrease in sperm motility was observed from 8 to 12 hours, indicating a significant decline compared to other adjacent time periods (p<0.05).
Discussion
During the process of sperm activation in bivalves, the amount of energy substances produced is relatively low. These energy substances include ATP and other metabolic by-products, which play important roles in the activation process.15 Additionally, some auxiliary substances may be produced, such as lactic acid and calcium ions, to maintain cellular environment stability and support sperm motility.17 However, compared to other biological processes, the amount of energy substances generated during sperm activation in bivalves is relatively low, which may be determined by their reproductive biology characteristics.24 When sperm and eggs meet in vitro, the sperm expend a significant amount of energy to adjust the osmotic pressure balance, leaving very little energy available for motility, hence the activated sperm exhibit a very short motility period.9 Studies have shown that sugars at appropriate concentrations can enhance sperm vitality.25 In this study, the results indicated that the mean effects on sperm vitality of appropriate concentrations of fructose and glucose were significantly superior to other additives. Under various activation conditions, fructose and glucose demonstrated significant differences in their impact on sperm vitality, effectively maintaining vitality and extending sperm lifespan. Among them, glucose showed a significant enhancement in sperm vitality compared to the control group, followed by fructose, this aligns with previous experimental outcomes.26 Thus, under different activation conditions, fructose and glucose as additives better extend and enhance the lifespan of Chinese pearl oyster sperm. This is because sugars like fructose, glucose, and galactose are important components of bivalve seminal plasma, participating in sperm metabolism and maintaining vitality. It has been reported that sperm are capable of tricarboxylic acid cycle metabolism, obtaining energy compensation through the oxidation of exogenous carbohydrates like glucose and fructose.27,28 Research shows that sperm have a high absorption efficiency for sugars such as fructose, glucose, and galactose, which are selectable energy sources for sperm activity and survival.29 These monosaccharides effectively extend the motility time of sperm, increasing the chances of contact between sperm and egg, thus significantly improving the fertilization rate in artificial insemination.30 Although both fructose and glucose are monosaccharides, their effects on the vitality of Chinese pearl oyster sperm are not completely identical, closely related to the energy metabolism efficiency of the two sugar solutions. The glucose solution is better at extending the vigorous motility time and lifespan of Chinese pearl oyster sperm than the fructose solution, making glucose more suitable as an activating solution for Chinese pearl oyster sperm.31 However, the specific mechanisms of absorption and utilization of these two monosaccharides by Chinese pearl oyster sperm warrant further study.
Sperm are highly sensitive to temperature variations, and their quality may be affected after exposure to different temperature environments during the production process.32 The results of this experiment indicate that sperm vitality is significantly better at low temperatures than at room temperature when fructose and glucose are added. This is because an appropriate temperature is necessary to promote sperm motility. Although high temperatures may accelerate the glycolysis of fructose and glucose, they can also irreversibly inhibit sperm activity or even lead to premature sperm death. Low temperatures suppress sperm motility, but they also reduce ATP consumption by sperm, lowering energy expenditure and thus extending sperm lifespan. In this experiment, the sperm retention time of fructose and glucose was prolonged by 8 hours at low temperatures compared to room temperature. This is consistent with previous research results.33 The sperm of Chinese pearl oysters adjust the lipid composition and fluidity of the cell membrane to enhance stability, optimize energy production and utilization to maintain life activities and regulate ion channels and transport proteins to maintain ion balance under low temperatures. Additionally, lowering the metabolic rate is more favorable for sperm survival.34 Cellular respiration and energy metabolism generally intensify with an increase in temperature, and the rapid consumption of intracellular energy shortens the lifespan of sperm. During the ex-situ preservation of sperm, it is advisable to control the environmental temperature within a reasonable range to low levels, thereby inhibiting sperm energy consumption and extending the ex-situ lifespan of sperm.29 Studies have shown that the lifespan, vitality, and fertilization capacity of sperm are closely related to the temperature of their environment.35,36 According to report,37 the short-term ex-situ preservation temperature for sperm is generally 4 °C, at this temperature sperm can move rapidly and their preservation time is extended. This demonstrates that adding fructose and glucose as exogenous energy sources to Chinese pearl oyster sperm under low temperatures can extend their lifespan. The vigorous movement time of sperm tends to decrease with increasing temperature, which is similar to the findings of most other sperm studies.
The reproductive biology of Chinese pearl oysters exhibits unique characteristics, where the sperm collected through artificial dissection remains inactive in both freshwater and seawater. This indicates that osmotic pressure does not fully initiate ion exchange in the sperm, thus requiring specific substances to thoroughly activate its ion channels. Ammoniated seawater, with a certain concentration of ammonia, is commonly used in production to activate the sperm.38 This experiment investigated the impact of ammoniated water on the vitality of Chinese pearl oyster sperm by comparing samples with and without the addition of ammoniated water. The results showed that under low-temperature conditions, the sperm vitality with added fructose and glucose was significantly better when ammoniated water was added compared to when it was not. Studies indicate that NH4+ can significantly affect sperm cells, being the only cation capable of opening ion channels in the membrane of Chinese pearl oyster sperm cells. It can fully open the ion channels on the membrane of these sperm cells, leading to an exchange of Na+ and Cl-. Consequently, the presence of NH4+ can extend the survival time of the sperm for several hours.39 Specifically, the impact of adding ammoniated water on sperm vitality is extremely significant within the first 0-24 hours of activation, with Chinese pearl oyster sperm showing a clear advantage when activated with ammoniated water compared to without. This is due to the volatilization effect of NH4+, which leads to the opening of ion channels. Additionally, adding ammoniated water helps maintain the structural stability of fructose and glucose, slowing down their decomposition rate, it has been reported that the activation of Chinese pearl oyster sperm relies on the presence of NH4+, which can maintain the ion channels in an open state.40 The addition of NH4+ may lead to changes in dissolved oxygen and pH in seawater, where the pH variation in seawater may affect the optimal sperm vitality and longevity.34 Future research should consider this factor and further explore the impact mechanism of NH4+ addition on sperm.
This study reveals that the factors of adding ammonia water, temperature, and additives each start to cause differences in sperm vitality after 0, 12, and 4 hours of activation, continuing until the end of the experiment. Under conditions with added ammonia water, the activation by NH4+ is completed instantly, indicating that the activation effect of NH4+ is direct, consistent with previous research findings.31 12 hours after activation, the temperature difference has a very significant impact on sperm vitality, with sperm vitality under low temperatures significantly superior to room temperature. This indicates that the impact of low temperatures on sperm vitality is not instantaneous but a slow and gradual process. Additives start to cause significant differences in sperm vitality 4 hours after activation, possibly because the process from hydrolysis to participation in sperm energy metabolism by fructose and glucose takes time. The main reason for the upward trend in sperm vitality within 4 to 8 hours is the energy compensation effect of fructose and glucose reaching a threshold at this time.27 Meanwhile, studies have shown that the hydrolysis of monosaccharides depends on factors such as temperature, and seawater salinity.38 During this period, the vitality of sperm with added ammonia water was significantly higher than that without ammonia water, possibly because the addition of ammonia water provided a more suitable environment for activity. Additionally, the addition of ammonia water accelerated the glycolysis of monosaccharides, which is beneficial for compensating the energy requirements of sperm. Under the same conditions, low temperatures significantly reduce the volatilization efficiency of ammonia water. This implies that the combined effects of ammonia water and low temperatures create an optimal condition for extending sperm vitality by providing an appropriate biochemical environment and reducing the rate of ammonia water loss, thereby maintaining its beneficial effects for a longer duration.41
The experimental exploration into the different factors affecting the vitality of Chinese pearl oyster sperm over a short period has shown that fructose and glucose, when used as additives, are significantly superior to the control group and other additives. Furthermore, storing at 4°C can slow down the decomposition rate of fructose and glucose, thereby extending the duration of energy compensation for sperm. Additionally, the introduction of ammoniated water reduces the reactivity of fructose and glucose, which is beneficial for maintaining their stability during the decomposition process. This temperature-controlled environment, combined with the stabilizing effects of ammoniated water, creates optimal conditions for preserving sperm vitality by ensuring a steady supply of energy substrates and minimizing the premature breakdown of these crucial nutrients. These findings highlight the importance of selecting appropriate additives and environmental conditions to optimize sperm vitality, particularly for enhancing the efficiency of artificial breeding practices in aquaculture. Therefore, fructose and glucose as additives, under low temperatures and with added ammonia water, provide suitable conditions for the activation and preservation of Chinese pearl oyster sperm.
Acknowledgements
This study was funded by the Central Public-interest Scientific Institution Basal Research Fund, CAFS (2023TD58), the earmarked fund for CARS (CARS-49), Financial Fund of Ministry of Agriculture and Rural affairs of China (NHYYSWZZZYKZX2020).
Authors’ Contribution
Conceptualization: Minghao Li (Lead). Data curation: Minghao Li (Lead). Formal Analysis: Minghao Li (Lead). Investigation: Minghao Li (Lead). Software: Minghao Li (Lead). Validation: Minghao Li (Lead). Visualization: Minghao Li (Lead). Writing – original draft: Minghao Li (Lead). Writing – review & editing: Zhengyi Fu (Lead). Project administration: Rui Yang (Lead). Methodology: Gang Yu (Equal), Wenli Zhou (Equal), Zhenhua Ma (Equal). Resources: Gang Yu (Equal), Zhenhua Ma (Equal). Supervision: Gang Yu (Equal), Wenli Zhou (Equal), Zhenhua Ma (Equal). Funding acquisition: Zhenhua Ma (Lead).
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
The Chinese pearl oyster (Pinctada fucata) is not a vertebrate or regulated invertebrate, so no ethical conduct approval is required.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.