

Introduction
The 2024 edition of The State of World Fisheries and Aquaculture (SOFIA)1 reported that global fisheries and aquaculture production reached 223.2 million tonnes in 2022, reflecting a 4.4% increase compared to 2020. This total included 185.4 million tonnes of aquatic animals and 37.8 million tonnes of algae. This expansion results from advancements across multiple domains such as production methodologies, nutritional strategies, pathological interventions, and genetic enhancements. An indispensable focal point within fish nutrition and pathology pertains to intestinal health, encompassing aspects of microbiome composition, structural integrity, and immune functionality. The intestine is a pivotal organ facilitating both nutritional assimilation and immunological defense, attributed to its rich innervation and intricate connections with the endocrine system.2 The gastrointestinal tract of fish operates in a milieu continuously exposed to dietary components and microorganisms from the aquatic habitat, potentially perturbing intestinal homeostasis through factors like nutritional imbalances, pathogenic invasions, and stressors.3 The primary functions of the fish intestine include nutrient absorption, water, and ion uptake, concurrently serving as a barrier against invasive threats. These functions hinge on the integrity of the epithelial layer, cell junctions, microbiota, and immune responses,4 all of which can be modulated by the incorporation of functional feeds.5
Functional feeds are typically enriched with elevated concentrations of ingredients, including minerals, vitamins, or immunostimulants, aimed at enhancing the health status and growth performance of animals. This paradigm shift in animal husbandry practices has been propelled by a departure from the conventional use of antibiotics, driven by their adverse consequences on the environment, animal welfare, and consumer health. Within the realm of aquaculture, Medium-Chain Fatty Acids (MCFAs) have gained prominence due to their organic origin and provision of readily available energy to animals. Kuang et al.6 have explored the application of MCFA as an antibiotic supplement, showcasing its antimicrobial effects and its observed protective influence on intestinal microarchitecture, a phenomenon previously documented in pigs.7 Key constituents within the MCFA family encompass caproic acid, caprylic acid, and capric acid. The strategic incorporation of functional feeds fortified with specific bioactive compounds, such as MCFAs, aligns with contemporary objectives to optimize animal health while mitigating the ecological and health concerns associated with traditional antibiotic use in animal agriculture. However, the rapid absorption of MCFAs in the gastrointestinal tract poses a challenge to their effective delivery to the deeper segments of the intestine, limiting their ability to exert full beneficial effects.8 This lack of effect may be attributed to their rapid absorption in the upper digestive tract, hindering their full contribution to improved gut health.8
Gut microbial community of fish is denser than terrestrial animals, owing to the fact that aquatic environments harbor a substantial abundance of microbiota .9 Studies have shown that the inclusion of dietary MCFAs contributes to growth-promoting effects and positively influences the gut microbial community. This is accomplished by augmenting the relative abundance of beneficial lactic acid bacteria in gilthead sea bream.8
Glycerol monolaurate (GML), a monoglyceride derived from medium-chain fatty acids, has been utilized as a food additive in the feed industry, as approved by the United States Food and Drug Administration.10 It is now widely acknowledged that symbiotic gut bacteria exert a lasting influence on the host’s immune system and metabolism through interactions involving microbial cell components and gene products. Recent studies have reported that GML notably increases the carcass yield, feed intake, and body weight in the male Cobb 500 broilers.11
GML maintains stability as it passes through the gastrointestinal tract, exhibiting a prolonged residence time.7 This implies that GML could directly impact the gut microflora10 and exhibit substantial antimicrobial potential, Thus, fostering a favorable environment for the promotion of beneficial effects on gut health.8 This facilitates direct interaction with the gut microbiota, which plays a crucial role in host health and physiology, especially regarding metabolism and immune development.12 Numerous studies have been carried out to explore the potential effects of GML on animals. For example, GML has demonstrated antimicrobial properties by directly influencing gut microbiota.13 Notably, GML may also contribute to nutrient metabolism and absorption within the gastrointestinal tract.14 To date, GML supplementation has shown significant potential in enhancing flesh quality and productive performance in both poultry15 and aquaculture species.16 Nonetheless, there is still a significant gap in research regarding the effects of GML on aquatic animals. Limited research indicates that GML can significantly enhance the growth of Danio rerio,16 Pelodiscus sinensis,17 and Larimichthys croceus.18Furthermore, GML is crucial in facilitating fat metabolism and reducing fat accumulation in Salmo salar.19 These studies suggest that GML could serve as a promising feed additive for enhancing gene expression, intestinal histology, and intestinal health in the culture of black sea bream fish.
Black sea bream (Acanthopagrus schlegelii) holds substantial economic significance in aquaculture, particularly in China, Japan, and various regions across Asia.20 The fish is a very popular aquaculture and abundant species in Southeast Asia, well-suited for intensive farming due to its rapid growth rate, strong disease resistance, and ability to tolerate a wide range of environmental conditions.16 Despite its economic importance, there is a notable dearth of research specifically addressing the nutrient requirements of black sea bream.21 Furthermore, a comprehensive understanding of the dietary composition necessary for Acanthopagrus schlegelii is lacking. Therefore, targeted investigations are warranted to elucidate the precise nutritional demands and dietary preferences of this species to optimize its aquacultural practices and ensure sustainable production. The purpose of evaluating GML as a dietary supplement for enhancing liver antioxidant capacity and gut health of juvenile black sea bream intestine.
Materials and Methods
Formulation of experimental diets and analyzing their composition
Six iso-nitrogenous (41.5%), iso-energetic (19 kJ g-1) diets were prepared in accordance with the regulator (GML 0.0), According to the black sea bream nutritional requirements; food was enhanced with a serial-wise increase in the level of GML 0.01%, 0.02%, 0.04%, 0.08%, 0.16%; named as GML1 0.01, GML2 0.02, GML3 0.04, GML4 0.08 and GML5 0.16, respectively. GML materials were obtained from the University of South China. Fishmeal (FM), soybean meal (SBM), soy protein concentrate (SPC), and squid liver meal constituted the main protein sources, whereas α-starch, fish oil, maize oil, and soybean lecithin served as the primary lipid sources (Table 1).
Each diet was prepared individually and processed as follows: raw ingredients were first ground into fine powder, then according to the formula, ingredients were weighed accurately and then mixed manually for 5 min. The food mixer was provided with mixed materials for stirring. Maize, Fish oil, and Soy lecithin were mixed separately and then added to the food mixer with the other materials. Water was slowly poured into the mixture and homogenized. With the help of an extruder (Model HUARUI, Wuxi, China; HKJ-218) equipped with a 2.5mm matrix. Finally, pellets were made of the mixture; the pellets were then dried by keeping them at 23°C for 72 hours, sieved, packaged in air-tight containers, and preserved at -20°C.
Animal husbandry, experimental site and conditions
Young black-seabream individuals were acquired from the Marine Fisheries Research Institute of Zhejiang, China. The experimental trials took place at the Joint Laboratory of Nutrition and Feed for Marine Fish, located at Zhejiang Marine Fisheries Research Institute on Xixuan Island in Zhoushan.
Before the growth trial, an indoor concrete tank (10m x 4m x 2m) was used for temporary storage of fish, and fish were kept acclimatizing fortnightly after being fed with commercial feed on a daily basis. Thereafter, 360 equal size fingerlings of IBW (initial weight of body) (1.55 ± 0.05 g) were distributed to eighteen cylinder-shaped fiberglass tanks, randomly with 350 L capacity, keeping the stocking bulk 20 Fish/tanks. Triplicate tanks were provided with randomly given dietary treatments. The fish were fed three times daily (at 8:00 AM, 12:00 PM, and 4:00 PM) over a period of 8 weeks. Daily feed consumption was monitored and recorded. Water supply was treated as follows: sea water was pumped from the sea and then stored for 48 hrs in a sediment pool; after that, sea water was filtered in a sand pool before reaching each tank. Each flow-through tank received sand-filtered, aerated seawater at a flow rate of 2 Lmin-1. The fish were kept under a natural photoperiod, with seawater temperature, ammonia nitrogen, and salinity in the tanks maintained at 27 ± 1 °C, 0.02–0.04 mgL-1, and 28 ± 2 gL-1, respectively. The pH ranged from 8.1 to 8.3, and dissolved oxygen concentrations were maintained at ≥5.0 mgL-1 throughout the experiment using air stones for continuous aeration. Depending on the breeding environment, tanks were cleaned 1h after the last feeding. The fecal collection began in 6th week of the growth trial. At 6:00 A.M, fecal samples were collected on routine basis following Bureau, Harris, and Cho22 and keeping it preserved -20 °C.
Samples collection
After completion of 8 weeks feeding trial, all experimental fish were starved for 24 hours and anesthetized with MS-222 solution of 60 mg L-1 (Sigma Chemical Co.).
Intestinal organ was the main target sample during this study; intestine tissue was sampled as follow:
For Biochemical, immune, and antioxidant indicators, the liver samples were stored at -80 °C. The samples of three full empty intestines per tank of each fish were collected and preserved at -80 °C for gene expression analysis. Three intestinal organs per aquarium were cut into two parts (anterior and posterior). About 3 cm of each intestine part were cut, washed, and cleaned with chilled phosphate buffered saline: PBS (pH 7.0), for further observation and morphological measurements secure in 10% formalin. 3 cm of each intestine part were preserved in PBS along with glutaraldehyde (2.5%) followed by storage at -4°C, for SEM examination.
Biochemical parameters
Assay kits from Nanjing Jincheng Bioengineering Institute (Nanjing, Jiangsu Province, China) were utilized to analyze serum samples. The goal was to measure various parameters, including total protein (TP), superoxide dismutase (SOD), malondialdehyde (MDA), glutathione peroxidase (GSH-Px), catalase (CAT), antioxidant capacity (T-AOC), alanine transaminase (ALT), and aspartate aminotransferase (AST). The experimental procedures closely followed the manufacturer’s instructions, as described in prior studies.23
Intestinal histology
Application of scanning electron microscopy (SEM) for microstructural analysis
Previously preserved gut samples were cleaned in glutaraldehyde and 0.1Molar pbs with pH: 7.0, After-fixed with osmium tetroxide (OsO4; 1%) for an hour or two. With a categorized ethanol sequence: 30%, then at 50%, 70%, 80%, 90%, 95%, and finally at 100% pure ethanol. After dehydration, it was then transported into a mixture of isoamyl acetate and alcohol (1:1) for 30 min overnight, kept in pure isoamyl acetic acid. Prior to observation, the dehydration of samples was performed for 4-5 min in a critical point dryer (Hitachi-Model HCP-2) through liquid CO2 and coated with gold-palladium in a Hitachi Model E-1010 ion sputter. Using Hitachi Model TM-1000 all samples were observed with SEM. Microvilli density was measured: in SEM photomicrographs with a magnification of 35,000 times, 3 images per diet treatment were selected. In the picture, microvilli were recognized by their round shape. The amount of the microvilli was calculated in an area of 1μm2; for each picture 3 area were selected, using Image-Pro plus 6.0 (Media Cybernetics, Inc.) software.
Analysis of gene expression patterns
Isolation of RNA and cDNA synthesis procedure
RNA extraction was conducted via Trizol kit (Invitrogen, USA). Almost 80 mg of RNA samples were isolated from gut samples. The RNA quality was confirmed by running gel electrophoresis using 1.5% agarose. Subsequently, the first-strand cDNA that served as template for Rapid Amplification of cDNA, ends PCR was manufactured using SMARTTM RACE cDNA Amplification Kit (Clontech), and the template cDNA used for Quantitative Realtime PCR (qRT-PCR) was synthesized using Primer ScriptRM reagent Kit (TaKaRa Biotechnology Co. Ltd., Dalian, China), both kits were handle as per company’s instruction. In order to eradicate genomic DNA, the reaction was incubated using gDNA Eraser (TaKaRa Biotechnology Co. Ltd, Dalian, China)) at 42 °C for 5 min. cDNA veracity was confirmed by running gel electrophoresis (1.5% agarose gel).
PCR reaction
The 25 μL PCR reactions were performed, comprising each primer of 1 μL, 2 μL of cDNA product, 12.5 μL of SYBRPremix-Ex/TaqII (from TaKaRa Company of Biotechnology, Dalian, China). The RT-qPCR was programmed, firstly, at 95 °C for a half an hour for initial denaturation, then 40 reaction cycles: 95 °C (duration 5 s), 60 °C (duration 30 seconds), an extension of 95 °C (duration 15 seconds). At the end of each PCR reaction, the amplification PCR product was confirmed to perform the melting curve analysis.
By 2−ΔΔCT method (Schmittgen and Livak, 2008), gene expression levels of Insulin growth like factor-I (IGF-I); Growth hormone (GH); Interleukin-1 (IL-1); Transforming growth factor (TGF); Tumor necrosis factor receptor 1 (TNF-1); Cholecystokinin (CCK); claudin-1 (CLDN-1), caspase-3 (CAS-3) and G protein-coupled receptor 41 (GPR41), were determined and normalized with β-actin. Separately each sample was studied through RT-qPCR in triplicate. IGF-1, GH, IL-1, TGF, TNF-1, CCK, CLDN-1, CAS-3, and GPR41 genes were recovered from the site: http://www.ncbi.nlm.nih.gov, using Primer (5.0) for designing of primers as per Table 2.
Analysis of microbial community composition
DNA extraction, PCR amplification, and Illumina MiSeq sequencing were conducted in this study. Fecal samples (n = 4) served as the source for total genomic DNA, extracted using the commercially available E.Z.N.A. Stool DNA Kit (D4015, Omega, Inc., USA). The PCR reaction mixture (12.5 μL of Pusion Hot start flex 2X Master Mix, 2.5 μL of each primer, and 50 ng of template DNA) targeted the V4-V5 regions of the bacterial 16S rRNA gene, employing universal primers 338F (5’ ACTCCTAC GGGAGGCAGCAG 3’) and 806R (5’ GGACTACHVGGGTWTCTAAT 3’). Amplification products were resolved on a 2% agarose gel, and the desired fragments were recovered using the AxyPrep PCR Cleanup Kit. The resulting pooled library underwent Illumina MiSeq sequencing, utilizing the MiSeq Reagent Kit V3 (600 cycles) for paired-end sequencing (2 × 300 bp).
Statistical examination and analysis
The data were presented as mean ± standard deviation (SD). Statistical analysis was conducted using one-way ANOVA, followed by Tukey’s post-hoc test. A significance level of p < 0.05 was considered for determining statistically significant differences. All statistical analyses were performed using IBM SPSS 20.0 (IBM, Chicago, IL, USA).
Results
Biochemical, immune and antioxidant indicators in the liver
Immune and antioxidant parameters were measured in liver. The SOD, TP, GSH-Px, CAT, MDA, AST, ALT, and T-AOC showed no significant effects (p > 0. 05) in all treatments (Table 3).
Gene expression in liver
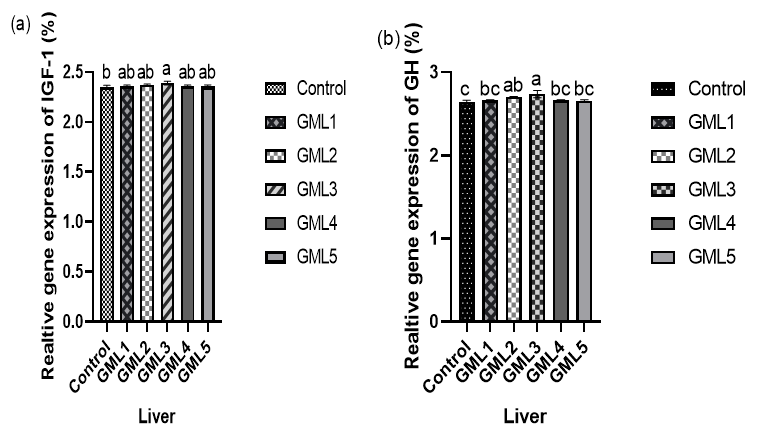
The Relative expression of genes, IGF-1 and GH genes were measured in liver. Relative gene expression in liver level of IGF-1 and GH were statistically higher in the GML3 treated group compared to other groups (p > 0.05) (Figure 1).
_insulin_growth_like_factor-i_(igf-1)_and_(b)_growth_hormone_(gh)_ge.png)
Intestinal gene expression
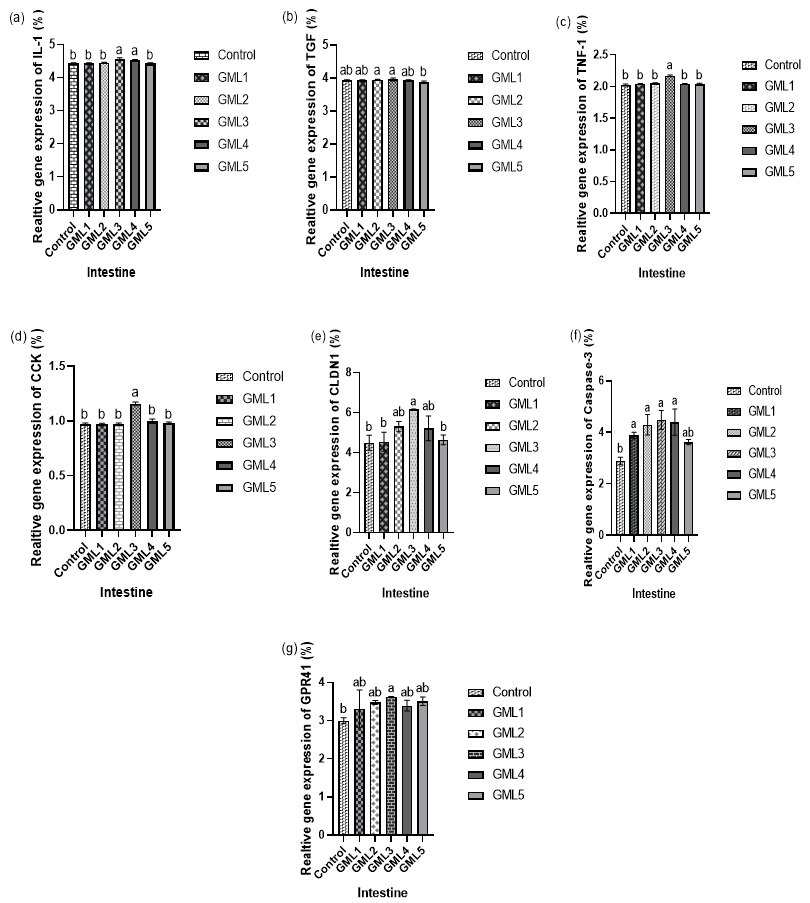
The relative expression levels of IL-I, TGF, TNF-1, CCK, CLDN1, CASS-3, and GPR41 genes were measured in the intestine, respectively. The relative expression of IL-I, TGF, TNF-1, CCK, CLDN1, CASS-3, and GPR41 genes was statistically higher (p > 0.05) in the GML3 group than in the other supplemented groups (Figure 2).
_interleukin-1(il-1)__(b)_transforming_growth_factor_(tgf)__(c)_tumo.png)
Intestine histological structure
The photomicrographs of Scanning electron microscopy (SEM) of the foregut and hindgut microstructure segment of black sea bream is shown in Figures 3,4. Fish from different dietary groups showed a gut with a columnar epithelium joined by good cells with great and systematized brush borders. The GML supplemented groups had longer brush borders compared to the control group. The cells seemed to be joined by an intact junctional complex without visible intercellular spaces.


Examining the composition and relative abundance of the microbiota community
Summary of sequencing data of intestinal microbiota
The sequencing data pertaining to the intestinal microbiota were presented in Table 4. In total, 1,251,273 raw tags underwent sequencing, yielding an average of 77,527.315 tags per sample, with individual counts ranging from 73,500 to 80,622. Following refinement, the dataset was reduced to 658,737 clean tags, averaging 71,360.557 per sample, with a range of 69,299 to 77,459. Taxonomic units of various bacterial species were subsequently identified from the clean tag pool, with all samples exhibiting Good’s coverage exceeding 0.999.
Alpha diversity and coverage estimate of intestinal microbiota
The dietary interventions did not significantly impact richness estimation (Chao1) and diversity measures (Simpson and Shannon indices). Across all six groups, 219 operational taxonomic units (OTUs) were identified within the intestinal microbiota. Notably, the GML3 fish group exhibited the greatest abundance of distinctive OTUs, while the control group exhibited the lowest (Table 5).
Phylum level
The composition of microbiota at the phylum level in the intestines of black sea bream was measured. A total of 8 phyla were identified in the fish subjected to various dietary treatments. Among these, Proteobacteria emerged as the most prevalent microbiota phylum across the groups, succeeded by Cyanobacteria, Firmicutes, Bacteroidetes, Acidobacteria, Actinobacteria, Verrucomicrobia, and Spirochaetes in sequential order. The phyla Bacteroidetes, Acidobacteria, and Verrucomicrobia showed no significant alterations (p > 0.05) in response to all dietary treatments. In general, the abundance of Proteobacteria, Cyanobacteria, Firmicutes, Actinobacteria, and Spirochaetes exhibited an increase in the treated groups compared to the control group (p < 0.05) (Table 6).
Class level
The microbiota composition at the class level was measured in different dietary groups. A total of 10 classes were identified in the intestines of fish subjected to dietary treatments. Gammaproteobacteria was the most abundant microbiota class among the groups, followed by Alphaproteobacteria, Oxyphotobacteria, Clostridia, Actinobacteria, Bacteroidia, Bacilli, Holophagae, Betaproteobacteria, and Negativicutes in that order. The Gammaproteobacteria, Alphaproteobacteria, Clostridia, Actinobacteria, Bacilli, and Betaproteobacteria of classes were not affected significantly, (p > 0.05) in all dietary groups. Overall, the Oxyphotobacteria, Bacteroidia, Holophagae and Negativicutes, increased with a GML supplementation (p < 0.05) (Table 7).
Genus level
The microbiota composition at the genus level were measured in different dietary groups. In the intestines of fish exposed to dietary treatments, a total of 20 genera were identified. The most abundant genera were: Chloroplast_unclassified > Pseudomonas > Delftia > Brevundimonas > Hydrogenophaga > Caulobacter > Sphingomonas > Novosphingobium > Mitochondria_unclassified > Pleomorphomonadaceae_unclassified > Alphaproteobacteria_unclassified > Achromobacter > Methyloversatilis > Dechloromonas > Stenotrophomonas > Lutispora > Clostridium_sensu_stricto_8> Subgroup_7_unclassified > Actinobacteria_unclassified > Curvibacter. The Pseudomonas, Hydrogenophaga, Caulobacter, Sphingomonas, Novosphingobium, Mitochondria_unclassified, Pleomorphomonadaceae_unclassified, Alphaproteobacteria_unclassified, Methyloversatilis, Dechloromonas, Stenotrophomonas, Lutispora, Clostridium_sensu_stricto_8, and Actinobacteria_unclassified of genera were not affected significantly, (p < 0.05) in all dietary groups. Overall, the Delftia, Brevundimonas, Achromobacter, and Subgroup_7_unclassified increased with a higher amount of GML supplementation (p < 0.05). While the Chloroplast_unclassified, and Curvibacter were reduced with a higher amount of GML supplementation (p< 0.05) (Table 8).
Discussion
GML is considered to have fungicidal, virucidal, anti-inflammatory and antibacterial properties.24 Previous reports suggests that the inclusion of antimicrobial drugs in the feed of poultry have several negative effects such as intestinal microbiota alterations, residues presence on meat and egg, eco-pollution, and leading to antibiotic-resistant microbes11Furthermore, the foremost consciousness of the health hazards to humans was activated by the absolutely excessive use of antibiotics in chicken feed, which caused an increase in the demand for studies to come up with a nature-based substitute. Presently, GML as a possible alternative was experimented with as an alternative for Zn-bacitracin. It was found that GML positively induced growth performance with the inclusion level 300 mgkg-1.
In this study, we examined the influence of GML supplementation on the well-being and nutritional absorption of black sea bream fingerlings. Various immune, biochemical, and antioxidant parameters were explored, including total protein (TP), superoxide dismutase (SOD), malondialdehyde (MDA), catalase (CAT), glutathione peroxidase (GSH-Px), alanine transaminase (ALT), total antioxidant capacity (T-AOC), and aspartate aminotransferase (AST). The findings revealed no significant impact of GML supplementation on the physiological and immune aspects of the treated groups of fish. Comparable results were observed in the previous study.23 The higher TP levels in the present study suggest improved protein metabolism.25 Furthermore, a decrease in MDA levels in the liver was observed, consistent with the findings of,26 who reported the antioxidant activities of malic and citric acids in their research on poultry.
In terms of biochemical parameters, AST and ALT exhibited no significant changes, indicating that GML exposure did not exert a substantial impact on the liver. Comparable outcomes were observed in previous studies involving lauric acid supplementation.23 Antioxidative enzymes were investigated in the present study to assess oxidative stress within the fish tissues. The overall antioxidant capacity is known as total antioxidant capacity (T-AOC). The study results revealed that GML impacted the activities of liver CAT and T-AOC content, indicating that GML has the potential to stimulate the production of antioxidant enzymes in the body, effectively counteracting reactive oxygen species (ROS). Our findings are consistent with the work of Seneviratne and Sudarshana Dissanayake.27 While limited data exist for aquatic animals, the literature supports our current findings: GML supplementation affected antioxidant parameters in black sea bream (Acanthopagrus schlegelii), which are consistent with observations by.23
Several genes play a crucial role in fish growth, with particular significance attributed to GHR and growth factor genes, such as insulin-like growth factor-1 (IGF-1).28 Growth factors are synthesized either via endocrine stimulation mediated by growth hormone (GH) or through paracrine/autocrine stimulation by the respective tissue. The growth hormone (GH)-insulin-like growth factor (IGF) axis functions as the principal pathway for growth and tissue proliferation in the majority of vertebrate species. In our current study, the expression of growth hormone receptor genes, such as GH, and growth factor genes, specifically insulin-like growth factor-1 (IGF-1), showed significant upregulation in response to GML supplementation. This observation aligns with earlier findings reported by.29 Research suggests that the fish liver is a primary site for insulin-like growth factor (IGF) synthesis, with secretion occurring in muscle cells as a paracrine hormone. The increased expression of IGF-1 RNA in liver tissues in our study supports this observation. Importantly, our study represents the first report on the modulation of growth-related genes, such as growth hormone (GH) and IGF-1, by GML in black sea bream.
In gram-positive bacteria the lipoteichoic acid (LTA) present in the cell wall and acts as a ligand for Toll-like receptor-2 (TLR-2), initiating a series of reactions involving cytokines such as tumor necrosis factor-1 (TNF-1) and interleukin-1 (IL-1).30 In black sea bream, GML supplementation was observed to enhance IL-1, TNF-1, and transforming growth factor (TGF). Existing literature indicates that diverse fish species can have their expression of proinflammatory cytokines, including IL-1, TNF-1, and TGF, effectively modulated by various diets.31 The study also noted an increase in proinflammatory cytokine genes expression like IL-1, TGF, and TNF-1 with probiotic supplementation. In our research, GML supplementation led to a significant upregulation of the anti-inflammatory cytokines IL-1, TGF, and TNF-1. Other studies have reported a relatively higher pro-inflammatory cytokine genes expression in tilapia post-feeding.32 Supporting this, tilapia exhibited an upregulation of IL-1 and TNF-1 gene expression, resulting in increased disease resistance.33 The observed upregulation of IL-1 gene expression following GML supplementation also suggests effective anti-inflammatory signaling.
This assertion is supported by a study that noted the upregulation of interleukin-1 (IL-1) in the intestine of O. niloticus due to feeding.34 Accordingly, our findings, which revealed significant increases in serum lipopolysaccharide (LPS) concentration and pro-inflammatory cytokines IL-1, transforming growth factor (TGF), and tumor necrosis factor-1 (TNF-1) in the GML group (Fig. 2 & 3), suggest that GML induces a notable state of systemic low-grade inflammation.
Cholecystokinin (CCK) plays a crucial role in regulating digestion and serves as a key factor in controlling appetite, contributing to the conclusion of meals in vertebrates. In the current investigation, there was a significant increase in CCK, aligning with earlier observations in the hindgut of ballan wrasse.35
Claudins, integral membrane proteins located in tight junctions of epithelial and endothelial tissues, play a crucial role in barrier function and inflammation regulation. In our study, CLDN-1 exhibited a significant increase, aligning with observations in a Northern American cohort.36
Extensive research has been conducted on caspase-3 genes in vertebrates, but the understanding of caspase functions in aquatic animals is still limited.37 The Caspase 3 family is recognized as a crucial immune gene in both vertebrates and invertebrates.38 However, due to the intricacies of fish tissue, a comprehensive understanding of how factors influencing apoptotic stimuli participate in apoptosis activation and affect mitochondrial mechanisms during postmortem fish storage remains elusive. Despite the thorough examination of the structure, characteristics, and functions of caspase-3 in various species, there is a scarcity of information regarding its role in oysters.38 This study unveiled an elevation in Caspase-3 levels with a GML-supplemented diet, consistent with similar findings in Eriocheir sinensis.39
GPR41, a G protein-coupled receptor (GPCR) found in mammals, is present in various human tissues such as peripheral blood mononuclear cells, adipocytes, and colon epithelial cells. The elevated expression of GPR41 in GML-supplemented groups suggests its potential significance in the intestine. Additionally, GPR41 has been identified in adipose tissue in mice and shown to stimulate leptin secretion.40
In animals, the villi of the intestine assume a crucial role in the digestion and absorption of food materials, concurrently fostering organismal immunity by providing a habitat for microbiota.41 It consists of epithelial cells that aid in the processes of digestion and absorption, with goblet cells generating mucus to safeguard underlying layers and provide lubrication for food materials. Intestinal integrity can be enhanced by dietary supplements like soybean meal,42 and studies suggest that medium-chain fatty acids (MCFAs), such as lauric acid, can serve as direct sources of energy for enterocytes.23 The current research is consistent with earlier results, suggesting that GML has a beneficial impact on the structural features of the intestinal mucosa in juvenile black sea bream. Comparable outcomes were observed in swine using MCFAs, where an increase in villi length in the small intestine, accompanied by reduced crypt depth and intraepithelial lymphocytes, was noted by Dierick et al.7 Furthermore, Hanczakowska, Szewczyk, and Okon43 reported that caprylic and capric acids, either provided individually or in combination, significantly enhanced piglet body weight and improved villus height.
The dynamic community within the intestines of fish consists of aerobic, partially anaerobic, and anaerobic bacteria, creating a unique and ever-changing environment known as the “island” microbiota. It is essential to preserve a harmonious intestinal microecology to support the optimal growth of fish, and achieving this balance depends on a diverse range of beneficial bacteria in the intestines. GML demonstrates a significant antibacterial impact, playing a role in stabilizing the balance of the animals’ intestinal microbiota.44
In this investigation, the alpha diversity indices (Simpson, Shannon, Chao1, and Observed species) of the black sea bream’s intestinal microbiota remained unaffected significantly. This suggests that the inclusion of GML did not exert a substantial influence on the overall diversity and richness of the microbiota. Nevertheless, when integrated into a high-lipid diet, the addition of GML resulted in a noteworthy augmentation of intestinal Shannon diversity in black sea bream. It is noteworthy that the reported significant impacts of GML in previous studies may have been influenced by nutrient manipulation, as demonstrated in the results of.45 Nevertheless, the GML groups in our study exhibited significantly higher abundance levels of Proteobacteria, Cyanobacteria, Firmicutes, Actinobacteria, and Spirochaetes at the phylum level, indicating the capacity of GML to modify the microbiota composition at this taxonomic level. At the phylum level, the predominant intestinal microbiota in juvenile T. ovatus consisted mainly of Proteobacteria, aligning with previous research findings.46 Key beneficial microflora, including Proteobacteria, Firmicutes, and Bacteroidetes, are recognized for supplying external digestive enzymes. These enzymes aid in breaking down food into smaller molecules, ultimately improving absorption and utilization in the fish gut.47 As highlighted by,48 up to 90% of the intestinal microbiota phyla in various fish species consist of Proteobacteria, Cyanobacteria, and Firmicutes. These phyla, including Bacteroidetes, Actinobacteria, and Verrucomicrobia, were among the top 8 phyla observed in black sea bream across all groups in our study. Prior investigations into the intestinal microbiota of black sea bream have consistently identified Proteobacteria, Cyanobacteria, Firmicutes, and Actinobacteria as dominant phyla,45,49 a pattern comparable to the top phyla observed in the control group in the present study. The higher abundance of Cyanobacteria and Spirochaetes in GML groups may indicate favorable conditions for their proliferation, contributing to a balanced microbiota diversity. While Spirochaetes were also identified as part of the dominant phyla in black sea bream by,49 their implications remain uncertain due to a lack of literature on the presence of this bacterial phylum in fish intestines. However, in our study, Proteobacteria, Cyanobacteria, Firmicutes, and Actinobacteria exhibited increased abundance in the treated groups compared to the control group. The higher abundance of Cyanobacteria and Spirochaetes in treated groups may have found the conditions created to be more favorable, warranting further studies to elucidate the significance of these bacterial phyla in fish intestinal microbiota. Our findings align with previous research in fish species, where Proteobacteria and Firmicutes were reported as the most dominant microbes in the fish gut, independent of the diet.50 The predominant phylum, Firmicutes, constituting approximately 70%, represents a transient microbial community in the distal section of the intestine.50 These bacteria are recognized as beneficial and are commonly employed as probiotics for vertebrates, particularly fish. In the present study, the addition of 0.04% GML was sufficient to augment the gut microbiota in the intestinal region of black sea bream, thereby promoting the proliferation of Firmicutes. Diets rich in plant materials are often associated with elevated levels of gut microbiota.50 Our findings indicate that dietary supplementation of GML led to a substantial increase in gut microbiota compared to the control diet, aligning with similar results reported by.23
However, in the present study, Oxyphotobacteria, Bacteroidia, Holophagae, and Negativicutes exhibited significantly higher abundance than the control group, indicating the capacity of GML to modulate the microbiota at the class level. The elevated number of Gammaproteobacteria suggests the utilization of plant-based diets and their involvement in cellulose degradation. The impact of the increased abundance of unclassified Oxyphotobacteria in the GML supplemented diet-fed fish remains unclear, primarily due to the lack of literature on its presence in fish. However, this class of Cyanobacteria is known to conduct oxygenic photosynthesis and has been detected in the microbiota of pig feces.51
Among the top 20 genera identified in the intestinal microbiota in this study, seven were significantly influenced by dietary interventions. Pseudomonas, Brevundimonas, and Novosphingobium were previously reported by45 as dominant genera in the intestinal microbiota of black sea bream. Conversely, the study by49 did not identify these genera as predominant in the same species. Remarkably, despite the observed pathogenic tendencies in certain Spirochaetes,52 the notably elevated presence of Brevinema (a Spirochaete) in the GML group cannot be attributed to pathogenicity, given the absence of disease symptoms in the fish. Healthy surgeonfish have exhibited a significant abundance of Brevinema in their intestinal microbiota.53 Therefore, it can be inferred that the increased abundance of Brevinema in fish fed the GML-supplemented diet may have contributed to the enhancement of intestinal structure and the expression of barrier protein genes. In our study, the abundance of Delftia in the intestines of fish from the GML-supplemented diet group could suggest that GML created favorable conditions for its proliferation. This finding aligns with previous research indicating the presence of this Proteobacteria genus in the intestines of various healthy fish, such as Atlantic salmon.54 Furthermore, additional studies are needed to substantiate this observation. Given the varied reports on intestinal microbiota composition and diversity, influenced by differences in the intestinal microbiome among various fish species and variations in rearing environments, reaching definitive conclusions about the implications or impacts of specific microbes may be paradoxical. This underscores the need for continuous research and meta-analysis in this field.
Our results align with previous observations regarding the impact of Lauric acid treatment on the gut microbiota. Both factors, Lauric acid treatment and alterations in the gut microbiota, exert influences on host health and physiology, particularly in aspects such as metabolism and immunological development.23
Conclusion
In essence, incorporating GML dietary supplementation into fish feed can markedly augment the liver immune index and antioxidant capacity of juvenile black sea bream. Our present findings underscore the considerable utility of GML3, with the 0.04% concentration proving optimal in our investigation. Additionally, supplementing the diet with GML3 (0.04%) leads to an increase in different genes and notable enhancements in both intestinal mucosal integrity and gut microbiota composition among juvenile black sea bream. Further elucidation of the precise mechanisms underlying GML’s effects warrants molecular-based investigations into the intrinsic connections between regulatory pathways and physiological functions.
Acknowledgments
This work was funded by the Key Research and Development Program of Ningbo, Grant Number: 2023Z113, and the Ministry of Science and Technology, China (Project No.:2020YFD0900801). We extend our gratitude to the China-Norwegian Joint Laboratory of Nutrition and Feed for Marine Fish situated on Xixuan Island in Zhoushan City, Zhejiang Province. Additionally, we thank the Key Lab of Mariculture and Breeding of Zhejiang Province for providing the experimental rearing system and logistical support during the fish growth trial.
Author contributions
Conceptualization: Sami Ullah (Equal), Fengqin Feng (Equal), Nanhai Xiao (Equal), Minjie Zhao (Equal), Jinzhi Zhang (Equal), Qingjun Shao (Equal). Data curation: Sami Ullah (Equal), Fei Shen (Equal), Jiahong Jian (Equal), Farman Ullah Dawar (Equal). Formal Analysis: Sami Ullah (Equal), Fei Shen (Equal), Nanhai Xiao (Equal), Jiahong Jian (Equal), Minjie Zhao (Equal), Farman Ullah Dawar (Equal), Jinzhi Zhang (Equal). Investigation: Sami Ullah (Equal), Fei Shen (Equal), Qingjun Shao (Equal). Methodology: Sami Ullah (Equal), Fengqin Feng (Equal), Qingjun Shao (Equal). Writing – original draft: Sami Ullah (Equal). Writing – review & editing: Sami Ullah (Equal), Fengqin Feng (Equal), Nanhai Xiao (Equal), Minjie Zhao (Equal), Farman Ullah Dawar (Equal), Jinzhi Zhang (Equal), Qingjun Shao (Equal). Supervision: Jinzhi Zhang (Equal), Qingjun Shao (Equal). Resources: Jinzhi Zhang (Equal), Qingjun Shao (Equal). Funding acquisition: Qingjun Shao (Lead).
Competing Interests
The authors of this manuscript disclose the absence of any conflicts of interest.
Ethical Statement
The experimental protocols employed in this investigation adhered to the Guidelines of the Care and Use of Laboratory Animals in China. Approval for the study was obtained from the Committee on the Ethics of Animal Experiments at Zhejiang University (Ethics code: ZJU20190052). Stringent measures were implemented to ensure the careful handling of all fish throughout the duration of the experiment.