



Introduction
Charybdis japonica and Portunus trituberculatus belong to the family Portunidae and are both economically valuable marine crabs.1 They are naturally distributed in the Yellow Sea, Bohai Sea, and East China Sea of China. The C. japonica is primarily harvested from wild natural resources, while the P. trituberculatus is the largest commercially farmed marine crab species in China. However, the farming yield of P. trituberculatus has been influenced by multiple factors such as climate, farming practices, and diseases.
Hemolymph in crustaceans serves as a primary mediator of both cellular and humoral immunity, with coagulation properties being indicative of its immune functionality.2,3 Previous studies have shown that various pathogens, including rickettsia, bacteria, fungi, and parasites, can cause coagulation abnormalities of hemolymph in crustaceans. Wang and coworkers4,5 found that rickettsia present in the hemolymph of trembling disease-infected E. sinensis could replicate the disease, and dissecting the diseased crabs revealed slow or absent coagulation of the hemolymph. He and coworkers6 discovered that Vibrio anguillarum and V. vulnificus could induce coagulation abnormalities and muscle opacity or whitening in P. trituberculatus. Yan and coworkers7 found that V. parahaemolyticus, V. alginolyticus, Pseudomonas aeruginosa, and other bacteria could cause autotomy of chelipeds and ambulatory legs in P. trituberculatus, with symptoms of slow or absent hemolymph coagulation and muscle opacity upon dissection. Fungi, including Candida oleophila,8 M. bicuspidata,9 and C. lusitaniae,10 have also been identified as causes of coagulation abnormalities in crustacean hemolymph. Xu et al. Isolated and confirmed the pathogenicity of the C. oleophila from the milky fluid in the carapace of “milky disease” affected P. trituberculatus. Similarly, Xu and coworkers9 found that M. bicuspidata could cause “milky disease” in E. sinensis. Wang and coworkers11,12 reported that infection with Hematodinium sp. in P. trituberculatus resulted in copious fluid discharge with a cloudy, noncoagulating appearance. Furthermore, mixed infections with multiple pathogens have been identified as causes of coagulation abnormalities. Wang and coworkers10 found that “milky disease” in P. trituberculatus was caused by co-infection with V. alginolyticus and C. lusitaniae, while Stentiford and coworkers13 identified yeast-like organisms co-infecting Necora puber and Cancer pagurus infected with Hematodinium sp. Although Zhao14 purified and detected white spot syndrome virus (WSSV) from tissues of artificially infected milky disease crabs, the disease symptoms were not reproduced in the infected crabs. From February to May 2022, we observed a sustained occurrence of hemolymph noncoagulation in C. japonica captured from the nearshore waters of Qinhuangdao. Although similar phenomena have been reported in cultured P. trituberculatus4 and cultured Eriocheir sinensis,15 scant literature exists regarding such pathologies in C. japonica. This study aims to explore the phenomenon of hemolymph coagulation abnormalities in C. japonica from a pathogenic perspective and identify potential pathogens.
C. japonica is a common species in nearshore areas, while P. trituberculatus is an economically significant aquaculture species along the coastal regions of China. There is a certain degree of commonality in the pathogens affecting crustaceans16,17, the larvae of C. japonica, harboring pathogens, may enter shrimp and crab aquaculture ponds through external seawater sources, potentially leading to disease outbreaks of P. trituberculatus. Therefore, we conducted infection trials and histopathological analysis on P. trituberculatus using dominant bacteria isolated from the hemolymph of C. japonica. Additionally, antimicrobial resistance and probiotic antagonism assays were performed on the isolated pathogens. The results of these experiments are expected to contribute to a better understanding of the sources and transmission pathways of pathogens causing diseases in P. trituberculatus, thereby providing theoretical support for the development of appropriate biosecurity measures and targeted disease control strategies.
Materials and Methods
Experimental animals and probiotic strains
The C. japonica was captured from the nearshore waters of Qinhuangdao, with an average weight of 100g ± 10g. A total of 20 individuals were collected. The crabs exhibited low vitality and showed no apparent signs of illness upon visual inspection. Following hemolymph extraction, the hemolymph failed to coagulate under ambient temperature conditions.
The healthy P. trituberculatus was sourced from a crab aquaculture facility located in Nanpai River, Huanghua. 80 of the P. trituberculatus had an average weight of 30g ± 3g, and were confirmed negative for specific pathogens through testing before being utilized in artificial infection experiments.
Probiotics: Bacillus mucilaginosus, Brevibacillus laterosporus, Lactobacillus plantarum, Enterococcus faecalis, B. subtilis, B. aryabhattai, and B. amyloliquefaciens were stored in the laboratory of the College of Marine Sciences, Hebei Agricultural University. B. paramycoides, B. toyonensis, and B. velezensis were stored in the laboratory of the Beidaihe Experimental Station, North China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences.
The main instruments and reagents
Microscope (Leica, DM4 B), McFarland Turbidimeter (Hangzhou Qiwei Instrument Co., Ltd., WGZ-XT desktop). Bacterial genome and yeast genome DNA extraction kits were purchased from Tiangen Biotech (Beijing) Co., Ltd., Loeffler’s alkaline methylene blue staining solution (0.1%), Gram staining solution purchased from Beijing Solarbio Technology Co., Ltd., LB liquid medium, 2216E medium, PDA medium, MHA medium, and other culture media were purchased from Guangdong Huan Kai Microbial Technology Co., Ltd., physiological and biochemical identification kits, antibiotic susceptibility test paper purchased from Hangzhou Tianhe Microbial Reagent Co., Ltd.
Initial inspection of diseased crabs
Referring to the method in Wang’s paper,18 the hemolymph from the pericardial cavity of C. japonica was aspirated using sterile syringes and subjected to smear examination under a microscope. Gill filaments were extracted and placed on slides for examination by immersion in water. The hepatopancreas were squashed by two slides for microscopic examination. The specimens were observed to ascertain the presence of parasitic or fungal infections.
Isolation and purification of bacterial strains
Hemolymph extracted from C. japonica was streaked onto 2216E agar plates and PDA agar plates. Following incubation at 30°C for 24 hours, prominent single colonies were selected for successive rounds of sub-culturing and purification, performed twice, to obtain pure cultures.
Morphological characteristics observation
The morphological characteristics of purified colonies were observed and recorded. Gram staining was performed on single colonies grown on 2216E agar plates, while methylene blue staining was conducted on single colonies grown on PDA agar plates.
Molecular biological identification
The total DNA of isolated strains was extracted using bacterial genomic DNA extraction kits or yeast genomic DNA extraction kits. Subsequently, the sequencing of bacterial 16S rDNA and fungal ITS sequences was outsourced to Sangon Biotech (Shanghai) Co., Ltd. The obtained sequences were compared for homology with the GenBank database. Based on the alignment results, sequences corresponding to strains with higher homology were selected, and a phylogenetic tree was constructed using the Neighbor-Joining (N-J) method implemented in MEGA7.0.
Physiological and biochemical characteristics identification
The purified strains were separately inoculated into physiological and biochemical identification tubes for the determination of physiological and biochemical indices. Reference was made to the “Manual of Common Bacterial Systematic Identification” and the “Manual of Characteristics and Identification of Yeasts”19 for preliminary identification of the isolated strains.
Drug susceptibility tests
Disk diffusion tests were employed for antimicrobial susceptibility testing. A volume of 100 µL of pathogenic bacterial suspension with a concentration of 108 CFU/mL was spread onto the surface of Mueller-Hinton agar (MHA) plates. After air-drying at room temperature for 3 minutes, 30 kinds of antimicrobial disks were placed onto the plates, with blank disks serving as negative controls. Each kind of antimicrobial disk was tested in triplicate, and the plates were incubated at a constant temperature of 30°C in an inverted position for 16 to 18 hours. The diameters of the inhibition zones were measured and averaged. The interpretation of the inhibition zone diameters was conducted according to the standard provided by Hangzhou Microbial Reagent Company’s “Interpretation Criteria for Antimicrobial Susceptibility Testing by Disk Diffusion Method” to determine the drug sensitivity of the pathogenic strains.
Determination of the antagonistic effect of probiotics on the isolated bacteria
The probiotic information including species, sources/strains, culture medium and culture temperature are given in Table 1. Ten probiotics were individually inoculated to reach the logarithmic growth phase. The bacterial cultures were then harvested and washed with 0.85% NaCl solution. Subsequently, the bacterial suspensions were adjusted to a density of 0.5 McFarland standard (approximately 1.5×108 CFU/mL) and further diluted to concentrations of 108, 107, and 106 CFU/mL using the same method. Similarly, pathogenic bacterial cultures were adjusted to a concentration of 108 CFU/mL using the aforementioned procedure for subsequent experiments.20
The agar dilution tests21 were employed to conduct antimicrobial susceptibility testing. A volume of 100 µL of pathogenic bacterial suspension with a concentration of 108 CFU/mL was evenly spread onto nutrient agar (NA) plates. After air-drying at room temperature for 3 minutes, sterile agar punchers (6 mm) were used to create wells with a minimum spacing of 2 cm between each well. Subsequently, 100 µL of probiotic suspension was added to each well. Three replicate wells were prepared for each concentration of probiotics. The plates were then incubated at 30°C for 18 hours in an upright position. The diameters of the inhibition zones were measured, and the average diameter was calculated.
Artificial infection test and histopathological observation
After a 7-day acclimation period, 80 healthy P. trituberculatus, with an average weight of 30g ± 3g, exhibiting smooth body surfaces and robust vitality, were selected for artificial infection. The test groups and injection conditions were shown in Table 2. The crabs were divided into four groups (three tests and one negative control), each consisting of 20 individuals. The experiment was conducted in a 200 L fiberglass tank with continuous aeration. For the test groups, a volume of 100 µL of pathogenic bacterial suspension with a concentration of 1×108 CFU/mL22 was injected into the muscle of each crab using a syringe. The control group received an equal volume of 0.85% NaCl solution by injection. Throughout the infection trial, the water temperature remained at 22°C, with a constant flow-through circulation system and no feeding. Crab behavior and mortality rates were observed and recorded every 24 hours. Hemolymph from moribund crabs were collected for pathogen reisolation. The experiment lasted for 120 hours. Additionally, gill filaments, hepatopancreas, chelae, and muscle tissues were fixed in 4% paraformaldehyde, sectioned, stained with hematoxylin and eosin (HE), and observed under a microscope.
Results
Symptoms and initial examination of crab disease
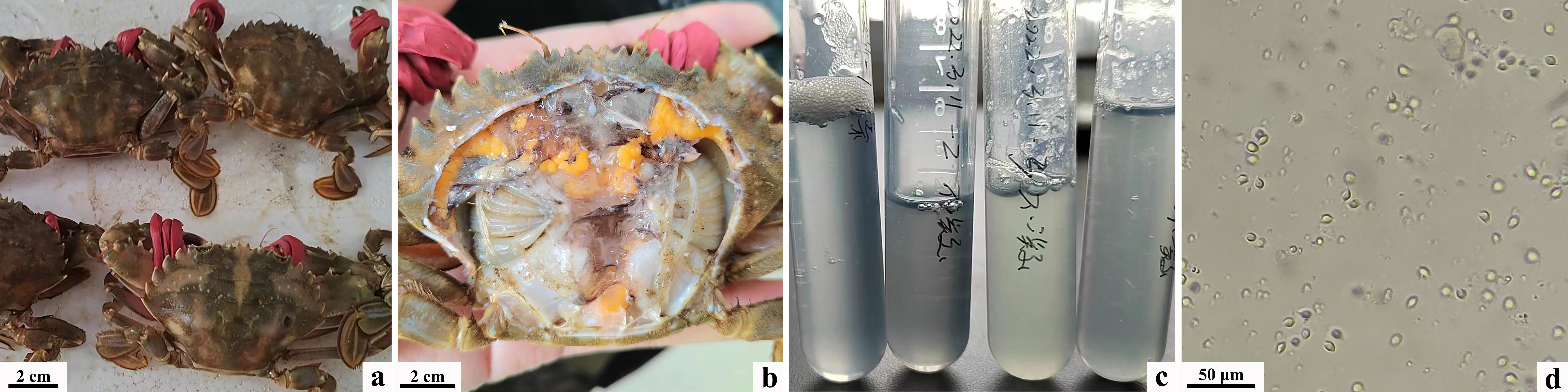
Wild-caught C. japonica exhibited poor activity, but did not show obvious external signs of disease. Upon dissection of the dorsal carapace, gill, hepatopancreas, and cardiac stomach tissues appeared normal in color. Hemolymph extracted from the pericardial cavity displayed a transparent bluish-green hue and failed to coagulate normally at room temperature. Examination under an optical microscope revealed an extremely low count of blood cells, with no evidence of blood flukes, microsporidia, or other parasitic organisms, as well as the absence of fungal hyphae or bodies (Figure 1).

Isolation, purification, and staining of pathogens
Under sterile conditions, hemolymph from diseased crabs was streaked onto 2216E agar plates and PDA plates. Two dominant bacterial strains were isolated from the 2216E plates and named N1 and N2, while one dominant bacterial strain was isolated from the PDA plates and named Q1. Colonies of strain N1 exhibited round, smooth morphology with well-defined edges, a raised center, semi-transparency, and a pale yellow color, characteristic of Gram-negative bacilli as observed in Gram staining. Colonies of strain N2 appeared round, milky white, and opaque, also exhibiting Gram-negative bacilli morphology. Colonies of strain Q1 displayed smooth edges, slight elevation in the center, and a white color, with observation after Loeffler’s methylene blue staining revealing varied sizes and morphologies indicative of budding reproduction (Figure 2).

Molecular biology identification
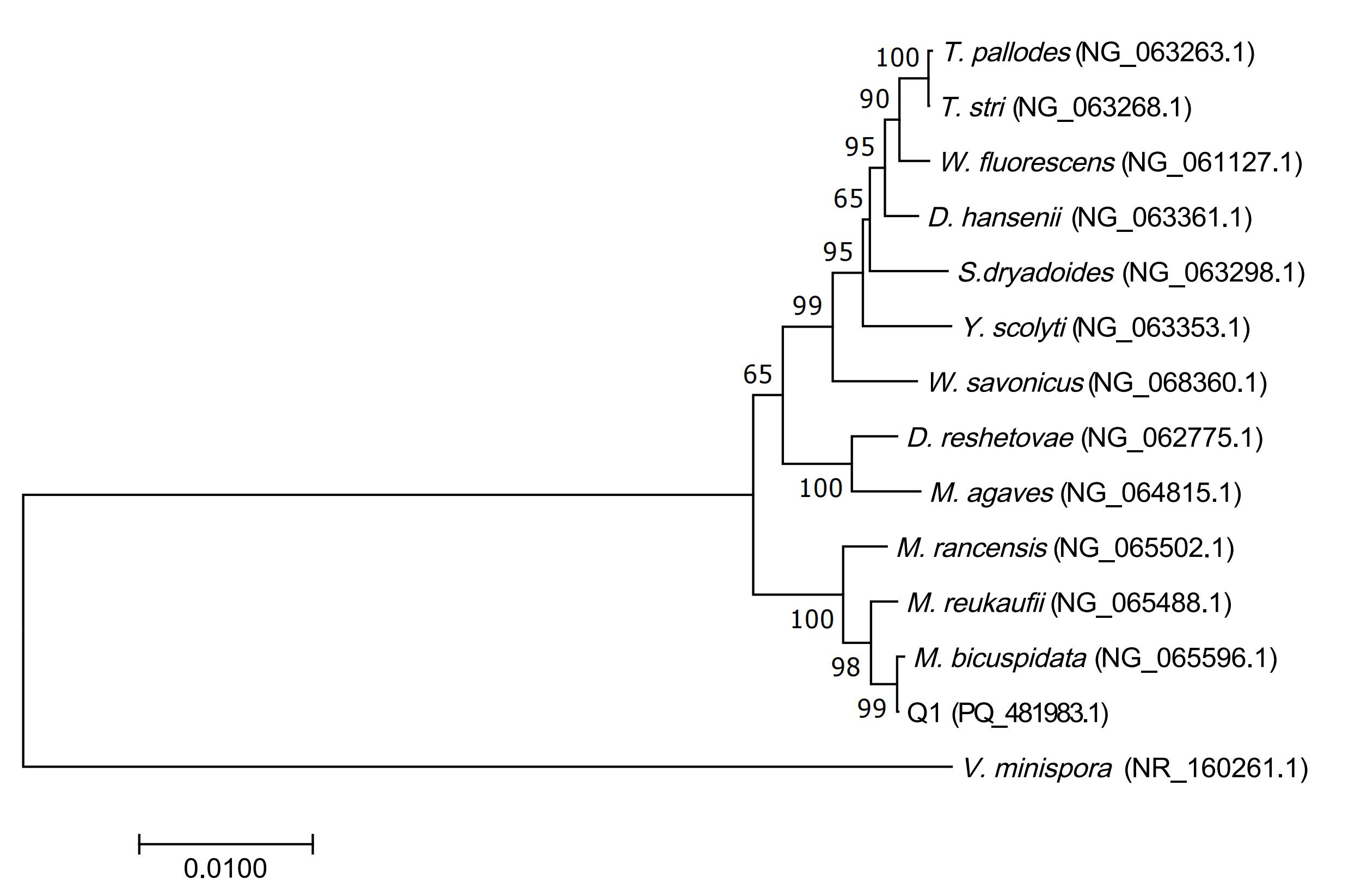
The obtained 16S rDNA sequences of the pathogenic bacteria N1 and N2, as well as the ITS gene sequence of Q1, were submitted to the GenBank database for sequence alignment. The results revealed that the pathogenic bacteria N1 (PQ_434666.1) shared 99.86% sequence identity with V. natriegens (NR_117890.1), N2 (PQ_434665.1) shared 99.37% sequence identity with V. fortis (NR_025575.1), and Q1 (PQ_481983.1) shared 99.64% sequence identity with M. bicuspidata (NG_065596.1). The Neighbor-Joining (N-J) phylogenetic tree constructed using MEGA7.0 showed that N1 and N2 belong to the Vibrio genus. N1 was most closely related to V. natriegens, forming a distinct branch, while N2 was most closely related to V. fortis, also forming a separate branch. Q1 clustered with M. bicuspidata, forming a distinct branch (Figure 3 & Figure 4).


Identification of physiological and biochemical characteristics
Pathogenic bacterium N1 was found to utilize sucrose, mannitol, and salicylic acid (glycoside), but not lysine, arginine, or ornithine. Conversely, the pathogenic bacterium N2 utilized lysine, arginine, and ornithine, but not amino acids or Citrate (Simon’s). Pathogenic bacterium Q1 utilized sucrose, sorbitol, and trehalose, while unable to utilize arabinose, rhamnose, or melibiose. The physiological and biochemical characteristics of N1 and N2 were consistent with those of V. natriegens and V. fortis, respectively, as described in the Bergey’s Manual of Systematics of Archaea and Bacteria. The characteristics of Q1 were in accordance with those of M. bicuspidata as outlined in the Handbook of Yeast: Characteristics and Identification.
Based on the morphological characteristics of the strains, molecular biology identification results, and physiological and biochemical characteristics, the pathogenic bacteria N1, N2, and Q1 isolated from the anomalous hemolymph of C. japonica were identified as V. natriegens, V. fortis, and M. bicuspidata, respectively (Table 3 & Table 4).
Drug sensitivity test results
According to the interpretive criteria for the zone diameters of antimicrobial susceptibility testing disks outlined in the clinical and laboratory standards institute guidelines, sensitivity testing was conducted for 30 antimicrobial agents against isolated strains N1, N2, and Q1. The inhibition zone diameters were measured and recorded, and the mean values were calculated. N1 and N2 exhibited sensitivity to antibiotics such as flomoxef and kanamycin (19-21 agents), while showing resistance to the remaining antibiotics. Q1 displayed sensitivity only to polymyxin B and exhibited resistance to the other 29 antimicrobial agents (Table 5).
Probiotic screening
Three isolated bacterial strains were subjected to antagonistic assays against 10 probiotic strains including B. mucilaginosus, L. plantarum, and B. subtilis. None of the 10 probiotic strains exhibited antagonistic effects against N1. B. velezensis, B. amyloliquefaciens, B. paramycoides, and B. toyonensis showed certain antagonistic activity against N2, with inhibition zone diameters decreasing as the probiotic concentration decreased. Six probiotic strains including B. velezensis, B. amyloliquefaciens, and B. mucilaginosus exhibited some degree of antagonism against Q1. Among them, B. amyloliquefaciens at a concentration of 107 CFU/mL showed the strongest antagonistic effect, with an average inhibition zone diameter of 23.6 mm, followed by B. amyloliquefaciens at a concentration of 108 CFU/mL, with an average inhibition zone diameter of 23.2 mm. B. mucilaginosus, B. laterosporus, and B. subtilis only exhibited inhibition at a concentration of 108 CFU/mL, with average inhibition zone diameters of 11.3 mm, 21.1 mm, and 14.5 mm, respectively (Table 6).
Pathogenicity analysis of the pathogen on the P. trituberculatus
The isolated bacterial strains N1, N2, and Q1 were each inoculated into healthy P. trituberculatus via intramuscular injection. Signs of mortality were observed in all experimental groups within 24 hours, and by the end of the 120-hour trial period, cumulative mortality rates for the N1, N2, and Q1 experimental groups reached 100%, 75%, and 35%, respectively, as summarized in Table 7. Upon injection with N1 or N2, the moribund crabs exhibited no apparent symptoms other than non-coagulated hemolymph. In contrast, those injected with Q1 showed milky white hemolymph with signs of coagulation; however, there was no accumulation of milk-like fluid in the cephalothorax cavity. Bacterial isolation from the hemolymph of artificially infected crabs yielded strains morphologically and sizably identical to those isolated from naturally infected crabs, namely strains N1, N2, and Q1, with consistent cultural characteristics (Figure 5).

Following infection with strain N1, vacuolar degeneration and desquamation of hepatopancreatic epithelial cells were evident; partial disappearance of gill filaments, and lamellae along with swelling of epithelial cells were observed in some areas of the gills; however, the muscle tissue appeared normal. Upon infection with strain N2, vacuolar degeneration of hepatopancreatic epithelial cells and infiltration of inflammatory cells into the interstitial spaces of the hepatopancreatic tubules were observed; notable ischemia of the gills with partial lamellar rupture was evident; inflammatory cell aggregation was observed in the interstitial spaces of the cheliped muscles. Infection with strain Q1 resulted in increased inflammatory cell infiltration and extensive necrosis in the hepatopancreas; notable ischemia of the gills, along with partial lamellar rupture; however, the muscle tissue appeared normal (Figure 6).

Discussion
Pathogens and diseases
Following the thorough examination of morphological characteristics, molecular biological identification, and physiological and biochemical properties, this study successfully isolated three pathogenic strains from the hemolymph of wild-diseased C. japonica: V. natriegens and V. fortis, and M. bicuspidata. Prior to this study, there have been no reports associating V. natriegens and V. fortis with coagulation disorders in crab hemolymph. Wang and coworkers23 reported that V. natriegens and V. harveyi could induce red-leg disease in Fenneropenaeus chinensis, leading to mortality. Li and coworkers24 found that a certain extracellular toxin produced by V. natriegens could result in mortality in clams and peacock fish. Chang and coworkers25 confirmed that V. fortis was a pathogenic bacterium affecting the sea urchin Strongylocentrotus intermedius, causing symptoms such as purple-red spots on the shell and clotting in internal tissues. Additionally, V. fortis has been implicated in disease outbreaks in rainbow trout and can lead to a mortality rate of up to 100% in early larvae of Artemia salina.26 The infection experiments conducted in this study revealed that both V. natriegens and V. fortis can induce coagulation disorders in the hemolymph of P. trituberculatus, with a pale blue color consistent with the original sample collected. Moreover, they exhibit relatively high mortality rates, with V. natriegens resulting in 100% mortality within 120 hours and V. fortis causing 75% mortality. Certain pathogens may demonstrate increased virulence at higher concentrations. By selecting the highest bacterial liquid concentration for infection experiments, the pathogenicity of the pathogens on the animal model under extreme conditions can be evaluated, thereby gaining deeper insights into their pathological mechanisms. Selecting the highest bacterial liquid concentration ensures the observation of significant infection effects within a relatively short period, thereby enhancing the reliability of experimental results. Ma and coworkers27 found that M. bicuspidata can cause “milk disease” in P. trituberculatus and E. sinensis, characterized by whitening and easy shedding of the walking legs of infected crabs, the presence of white milky fluid in the cephalothorax, and generalized muscle edema with reduced plumpness. In contrast, the results of this experiment showed that the isolated Candida ortholacerationis can lead to a milky appearance in the hemolymph of C. japonica but did not result in coagulation disorders, and there were no typical symptoms of milk-like fluid accumulation in the cephalothoracic cavity observed in “milk disease”.
The significance of drug resistance analysis and probiotic screening for production guidance
The antimicrobial susceptibility analysis of the isolated V. natriegens in this study exhibited results similar to those obtained by Li and coworkers24 from the isolation of V. natriegens from the hepatopancreas of Crassostrea rivularis at a culture farm in Wenzhou, China. However, significant differences were observed in the sensitivity to antimicrobial agents such as compound trimethoprim, amikacin, and erythromycin. Previous research has indicated that differences in antimicrobial susceptibility among strains of the same species from different sources may be attributed to regional variations, laboratory culture conditions, and antibiotic usage practices.28 Nevertheless, there was a paucity of reports on the antimicrobial susceptibility analysis of V. fortis and M. bicuspidata. The resistance results of the three pathogenic strains revealed that ciprofloxacin and kanamycin exhibited the strongest inhibitory effects against V. natriegens and V. fortis, while polymyxin B displayed good inhibitory effects against all three pathogenic strains. These findings would contribute to the in-depth study of the pathogenic mechanism, and would help to develop rapid and accurate treatment strategies of the three isolated strains. Otherwise in this study, 18 drugs prohibited in aquaculture, including Minocycline、Azithromycin、Erythromycin、Levofloxacin、Norfloxacin、Chloramphenicol、Clindamycin、Trimethoprim、PolymyxinB、Ciprofloxacin、Penicillin、Ceftazidime、Cefperazone、Cefazolin、Cephalexin、Cefuroxime、Ceftriaxone and Vancomycin,were used to test the inhibitory effect of the isolated pathogens, and 14 of them had antibacterial effects on N1, N2 or Q1. Therefor the results would remind that the supervision of these drugs should be strengthen in practical production settings.
In the effort to reduce antibiotic usage, the use of probiotics to antagonize pathogens is a promising approach which deserves further exploring. Streptococcus agalactiae is a significant pathogen in the intensive culture of tilapia. Studies have shown that two strains of Bacillus isolated from Nile tilapia exhibit significant inhibitory effects against it.29 Lactic acid bacteria isolated from benthic organisms not only produce hydrolytic enzymes and secondary metabolites but also effectively reduce the number of V. parahaemolyticus, a common pathogen in aquaculture.30 In this study, B. velezensis and B. amyloliquefaciens at concentrations above 106 CFU/mL exhibited antagonistic effects against V. fortis and M. bicuspidata, while 10 strains of probiotics showed no antagonistic effects against V. natriegens. Further expansion of the range of probiotic selection is needed. This result also suggests that the work of selecting probiotics with antagonistic effects against aquatic pathogens in aquaculture needs to be further strengthened.
Conclusion
In summary, this study represents the first isolation of V. natriegens, V. fortis, and M. bicuspidata from diseased C. japonica. Moreover, all three pathogenic strains demonstrated pathogenicity towards P. trituberculatus. These findings contribute to a better understanding of the sources and transmission pathways of pathogens in cultured P. trituberculatus, facilitating the development of appropriate biosecurity measures. The investigation into the drug resistance of these three pathogens and the antagonistic effects of probiotics provides theoretical support for the precise prevention and control of aquatic animal diseases caused by these pathogens, as well as for the development of related biological products.
Acknowledgments
This research was supported by Hebei Modern Agricultural Industry System (HBCT2023220207), and Special Fund for Talent Introduction of Hebei Agricultural University, China (Grant Nos. YJ2019023).
Authors’ Contribution
CRediT taxonomy:
Conceptualization: Yawei Qin (Equal), Yanmiao Tan (Equal). Formal Analysis: Yawei Qin (Lead). Software: Yawei Qin (Lead). Validation: Yawei Qin (Lead). Visualization: Yawei Qin (Lead). Writing – original draft: Yawei Qin (Lead). Methodology: Yanmiao Tan (Equal), Chunguang Gong (Equal). Project administration: Yanmiao Tan (Equal), Chunguang Gong (Equal). Writing – review & editing: Yanmiao Tan (Equal), Chunguang Gong (Equal). Investigation: Yufeng Liu (Equal), Zishuo Bai (Equal), Jianmeng Wang (Equal). Resources: Yufeng Liu (Equal), Tianlei Yang (Equal). Data curation: Zhongwei He (Equal), Tian Han (Equal). Funding acquisition: Chunguang Gong (Lead). Supervision: Chunguang Gong (Lead).
Competing of Interest – COPE
The authors declare that they have NO competing financial interest exist.
Ethical Conduct Approval – IACUC
All experiments and handling of animals were carried out according to the research protocol approved by the Animal Care and Use Committee of Hebei Agricultural University and Beidaihe Central Experimental Station.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.