Introduction
The largemouth bass (Micropterus salmoides), or California bass, is a freshwater fish native to the Mississippi River basin in the USA and has been introduced to many regions worldwide.1 Largemouth bass has gained popularity for its delicious flesh, rapid growth, and strong adaptability to a variety of environments in China.2 Approximately 802,486 tons of M. salmoides were produced in China in 2022.3 However, the largemouth bass farming sector is increasingly vulnerable to disease threats due to the increasing aquaculture density.4,5 The largemouth bass rhabdovirus (LMBRaV) has emerged as one of these threats and has threatened largemouth bass breeding globally.6,7 LMBRaV is a polyhedral virus with a diameter of 120-300 microns, consisting primarily of single-molecule linear double-stranded DNA viruses with a genome size of 170-200 kb. The major capsid protein (MCP), constituting 40%-45% of the virus’s total protein content, exhibits highly conserved characteristics useful for distinguishing various ranaviruses.8 Previous studies have shown that some drugs, such as epigallocatechin-3-gallate (EGCG) and Saikosaponin C, have potential anti-LMBRaV and immunomodulatory activity.9,10 However, owing to concerns about cost and production yield, these drugs have not yet been extensively employed in aquaculture. Therefore, it is vital to develop effective strategies for managing and controlling LMBRaV.
Given the immune system’s pivotal role in defending aquatic animals against pathogens, enhancing host immune function is crucial for preventing disease outbreaks.11 In the innate immune response against viruses, type I interferons (IFNs) play a crucial role, while myxovirus resistance (Mx) proteins exert direct antiviral effects by blocking viral genome replication at an early stage.12 In addition, MHC-1 molecules facilitate antigen presentation to effector T cells in vertebrates during adaptive immunity.13 Immunomodulators, derived from natural and synthetic substances, offer promising avenues for regulating host immune function.14 In aquaculture, Chinese herbal medicine has long been utilized to modulate immune responses and avert the outbreak of diseases herbal medicine has long been utilized to modulate immune responses and avert the outbreak of diseases.15 Extensive research shows that components isolated from medicinal plants exhibit immunostimulant, growth-promoting, anti-stress, and disease-resistant effects in aquaculture.16 As an example, dihydroartemisinin and tetrandrine significantly inhibited spring carp viraemia of carp virus by activating innate immunity and reducing apoptosis.17 Additionally, researchers found that coumarin can boost the levels of IFN-related genes to counter infectious haematopoietic necrosis virus (IHNV) infection.18 In largemouth bass, piperine, Saikosaponin C and epigallocatechin-3-gallate showed promising antiviral and immunomodulatory effects.9,10,19 However, the high prices and low yields of these drugs hinder their extensive use in largemouth bass farming. Consequently, identifying herbal plant components with enhanced efficacy and better suited for aquaculture may offer an effective strategy to control LMBRaV infection in the future.
This study focused on the antiviral and immunoregulatory role of quercetin against LMBRaV infection by assessing antiviral stability, the degree of cell nuclear damage, and innate immune response experiments. Forthermore, we examined the effect of quercetin against LMBRaV infection in vivo through survival trials, immune-related gene expression, and histopathological observations. These findings offer theoretical insights for the development of novel antiviral drugs in aquaculture.
Materials and Methods
Cell, virus, and fish
The epithelioma papulosum cyprini (EPC) cell line was preserved in our laboratory. LMBRaV was isolated and identified from diseased largemouth bass and subsequently cultured in EPC cells until approximately 80% exhibited cytopathic effects (CPE). The infected cell culture containing LMBRaV was then subjected to three cycles of freeze-thawing. After that, virus fluid was gathered and divided into cryovials, which were kept for later use at -80°C. The TCID50 was determined using the Reed-Muench method.
Largemouth bass (body weight 7.5 ± 0.5 g) were procured from a fish farm in Wuhan, China. After testing, they were confirmed to be healthy and free of LMBRaV. Under laboratory conditions, the largemouth bass were acclimated for 14 days in 300 L recirculating aquariums. During acclimation, the fish were fed commercial feed (Tongwei, Chengdu, China) twice daily. The water temperature was maintained at 25°C ± 1°C. PCR analysis utilizing LMBRaV MCP gene primers (Table 1) confirmed the absence of LMBRaV in the largemouth bass.
Compounds cytotoxicity assay
EPC cells were cultured and seeded in a 96-well plate. M199 medium containing 5% FBS was used to dilute the stock solution to pre-defined concentrations. After 48 hours of incubation with the relevant drugs, the old culture media were aspirated, followed by the sequential addition of 100 μL fresh M199 culture medium and 5 μL CCK-8 (Beyotime, China) reagents. After 2 hours of incubation in the cell culture incubator, absorbance at 450 nm was measured using a microplate reader (ELX-800, China)
Anti-LMBRaV activities assay
EPC cells at a density of 1×105/mL were inoculated into a 12-well cell culture plate. When the cells converged to a monolayer, the cells were incubated with LMBRaV virus suspension at a concentration of 103 TCID50/mL at 25°C for 2 hours. After the incubation of LMBRaV, the virus solution was recovered, the EPC cells were rinsed with M199 for 2-3 times, the cell maintenance solution containing natural compounds was added. The diluted compounds solution were then added at the maximum safe concentration, and the resulting mixture was incubated at 25°C for 72 hours. Finally, the cells were digested and harvested, and total RNA of each sample was extracted. RT-qPCR was used to detected the LMBRaV MCP gene expression in order to assess the antiviral efficacy of compounds against LMBRaV.
Cell nucleus damage assay
EPC cells were inoculated in confocal petri dishes and cultured for 12-24 h, followed by infection the LMBRaV with a titer of 103 TCID50/mL for 2 h at 25°C. Subsequently, replaced with cell maintenance solution containing quercetin (20 mg/L) and incubated for 48 h. The cells were washed three times with 0.1 M PBS, fixed for 30 min in 4% paraformaldehyde, and then stained with 1 mg/L DAPI, 5 mg/mL Dil, staining. Fluorescence was observed with a confocal microscope (Olympus FV3000, Japan).
Effects of different treatments on LMBRav proliferation
Viral binding tests was assessed according to a previously established protocol,17 in which the virus binds selectively to the cell membrane rather than to internal organelles. A total RNA extract of the collected cells was then performed, and RT-qPCR was used to detect the expression of LMBRaV MCP gene to evaluate the viral binding.
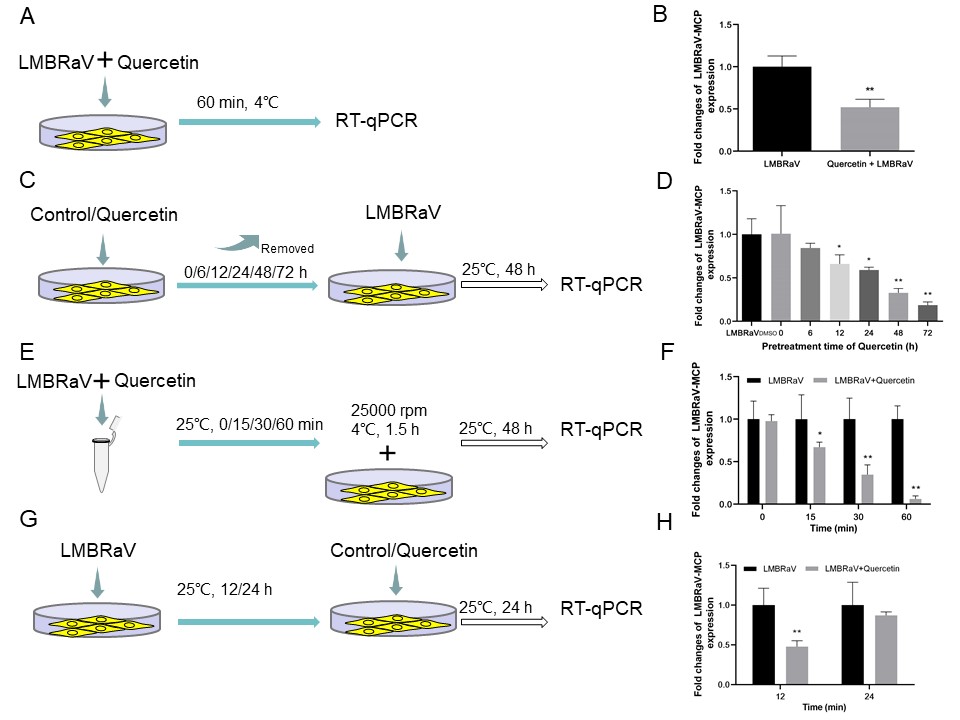
Pre-treatment of quercetin: EPC cells diluted to a certain density were inoculated in 12-well plates. When the EPC cells grew to a monolayer, the cells were rinsed with M199 for 2-3 times. And then the cells were exposed to quercetin (20 mg/L) at 25°C for 0, 6, 12, 24, 48, and 72 h. After incubation, the old solution was removed, and then inoculated with LMBRaV (103 TCID50) at 25°C for 2 h. The viral solution was then recovered and incubated in cell maintenance solution for 48 h. The cell samples were collected, and the total RNA of the samples was extracted, and then the expression of LMBRaV MCP gen was detected by RT-qPCR.
LMBRaV and quercetin co-incubation assay: LMBRaV (103 TCID50) viral solution were co-incubated for 0, 15, 30 and 60 min at 25°C with 20 mg/L quercetin. After incubation, the treated LMBRaV-quercetin mixture was added to 12-well plates grown to monolayers of cells, and the cells were collected after continuing to incubate the cells for 48 h at 25°C. Finally, the cells were digested and harvested, and total RNA of each sample was extracted. RT-qPCR was used to detected the LMBRaV MCP gene expression.
Quercetin Post-Processing: LMBRaV (103 TCID50) virus solution was added to 12-well plates grown to monolayers of cells, incubated at 25°C for 2 h. The virus solution was recovered, and cell maintenance solution was added to continue incubation for 12 or 24 h. The old solution was removed, and the cells were rinsed twice with M199, and cell maintenance solution containing quercetin (20 mg/L) was added, and the cell samples were collected after 24 h. The total RNA of each sample was extracted. RT-qPCR was used to detected the LMBRaV MCP gene expression.
Toxicity tests of quercetin in largemouth bass
In this research investigation, largemouth bass were administered varying doses of quercetin (5, 10, 20, 30, 50, and 70 mg/kg) in order to evaluate potential toxicity. Mortality rates among the largemouth bass were monitored and recorded on a daily basis over a period of 15 days.
The protective effects assays
In the survival rate assay, a cohort of healthy largemouth bass was randomly allocated into three experimental groups: PBS + DMSO, LMBRaV + DMSO, and LMBRaV + quercetin, with 30 fish per group and 3 replicates. LMBRaV + DMSO and LMBRaV + quercetin groups were intraperitoneally injected with LMBRaV diluent (103 TCID50, 100 µL/fish), Control groups received an equivalent volume of M199 via intraperitoneal injection. Diets containing quercetin (20 mg/kg) or DMSO equivalents were given to the fish after a 12-hour infection period. Subsequently, the fish were monitored at 24-hour intervals over a period of 10 days to document mortality.
The viral load assay was conducted on a cohort of healthy largemouth bass, stratified into two experimental groups: LMBRaV + DMSO and LMBRaV + quercetin. Specimens from each group were subjected to viral load assessment at 1, 3, and 5 days after LMBRaV infection.
The preventative effects assays
Fish were divided into three experimental groups: PBS + DMSO, LMBRaV + DMSO, and LMBRaV + quercetin, to assess whether quercetin prevented LMBRaV infection. For a period of five days, diets containing quercetin (20 mg/kg) or DMSO equivalents were given to the fish. Following this, LMBRaV (105 × TCID50/mL) was administered to the virus-infected groups, while M199 in a similar volume was administered to the control groups. In addition to the survival rate, fish tissues were obtained at 1, 3, and 5 days after LMBRaV infection for the evaluation of immune-related gene expression, the assessment of histopathology, and the analysis of viral load.
Histopathological examination
Three fish are processed after they have been infected with the LMBRaV for five days. The liver, spleen, and kidney are removed and fixed in 4% paraformaldehyde. The samples are then embedded in paraffin, sectioned, and stained with haematoxylin and eosin. Morphological changes in the liver, spleen and kidney are observed under the microscope.
Statistical analysis
The statistical analyses were performed using GraphPad Prism 8. A one-way ANOVA was used to evaluate the differences between groups. The differences were deemed statistically significant if the p-value was less than 0.05.
Results
Antiviral activities of natural compounds
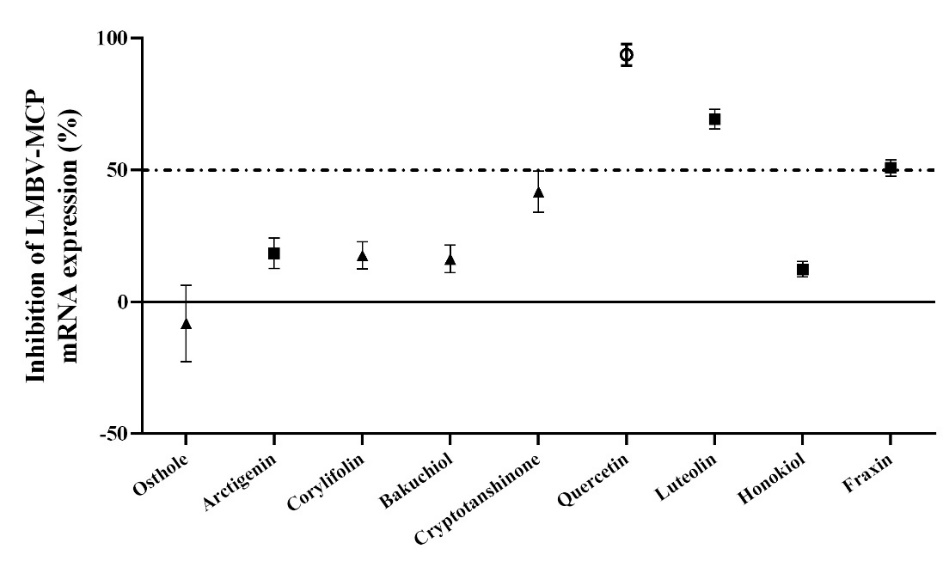
The inhibitory effects of nine compounds on LMBRaV were evaluated (Figure 1). Quercetin demonstrated the most potent anti-LMBRaV activity, suppressing LMBRaV-MCP gene expression by more than 93.6%. Consequently, we suggest prioritizing research on quercetin’s effectiveness against LMBRaV.

Antiviral activity of quercetin in vitro
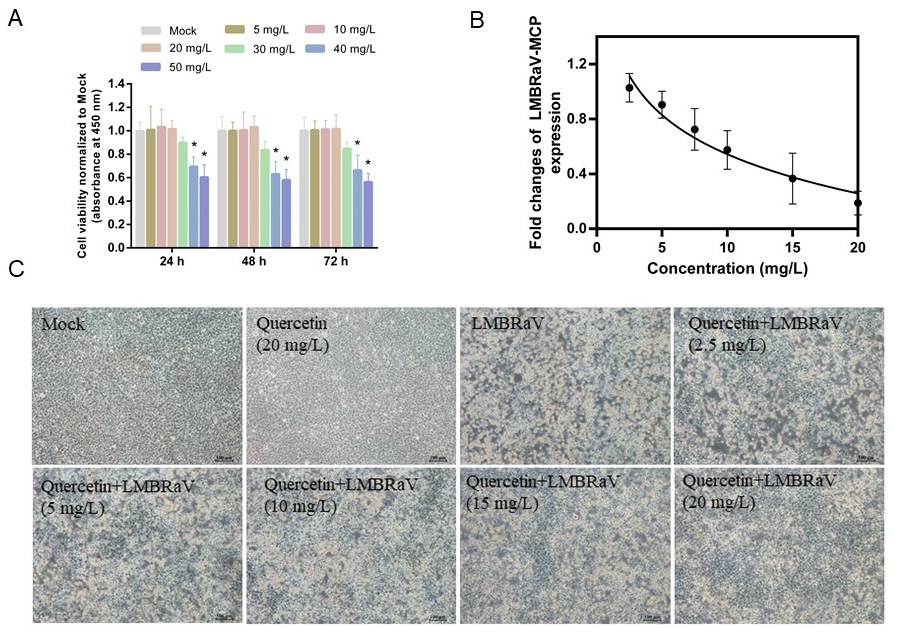
According to the cytotoxicity studies of quercetin, 20 mg/L of quercetin did not do significant harm to EPC cells for the duration of the experiment (Figure 2A). The most effective antiviral response was observed at 20 mg/L, where quercetin inhibited LMBRaV MCP gene expression dose-dependently (Figure 2B). The morphology of EPC cells infected with LMBRaV resembled fragmented fishing nets. However, quercetin significantly reduced the CPE phenomenon caused by LMBRaV infection (Figure 2C).
_effect_of_quercetin_at_different_concentrations_on_.jpeg)
Effect of quercetin on LMBRaV-induced nuclear damage
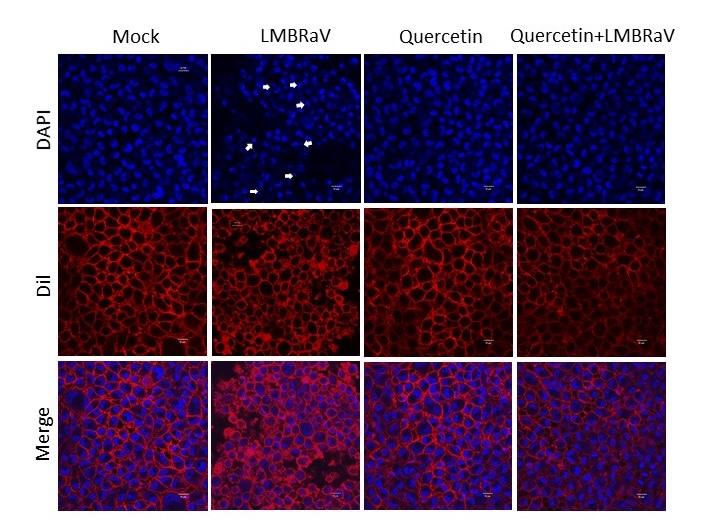
The cells were stained using DAPI/Dil staining solution to observe the nuclear damage caused by LMBRaV and the effect of quercetin treatment on the nucleus, and the results are shown in Figure 3. The results demonstrated that LMBRaV infection led to substantial nuclear damage in EPC cells. Treatment with quercetin resulted in decreased LMBRaV-induced nuclear fragmentation. This indicates that quercetin can protect the nucleus from damage and has a good anti-LMBRAV effect.

Effects of quercetin on viral infectivity
The results of the particle adsorption assay showed that quercetin effectively inhibits the adsorption of LMBRaV on EPC cells (Figure 4A). The results of LMBRaV proliferation in cells after quercetin pre-treatment of EPC are shown in Figure 4D. When quercetin was incubated for 12, 24, 48 and 72 h and then infected with LMBRaV was able to significantly reduce the virus proliferation in EPC cells. Figure 4E depicts an experimental investigation into the potential influence of quercetin on LMBRaV toxicity in M199. The subsequent Figure 4F illustrates that quercetin had a direct killing effect on LMBRaV, and quercetin had the best direct inactivation effect on the virus when the LMBRaV - quercetin was co-incubated for 60 min, and the intracellular LMBRaV MCP gene expression decreased by 93.89%. Another experiment detailed in Figure 4G examined the impact of quercetin post-treatment on LMBRaV replication. Notably, Figure 4H showed that quercetin still had significant antiviral activity 12 h after LMBRaV infection of EPC cells. Compared with the control group, the LMBRaV MCP gene expression within EPC decreased by 51.14% after quercetin treatment, respectively. However, when LMBRaV infected EPC cells for 24 h, the cells treated with quercetin had no anti-LMBRaV effect.
_the_virus_.jpeg)
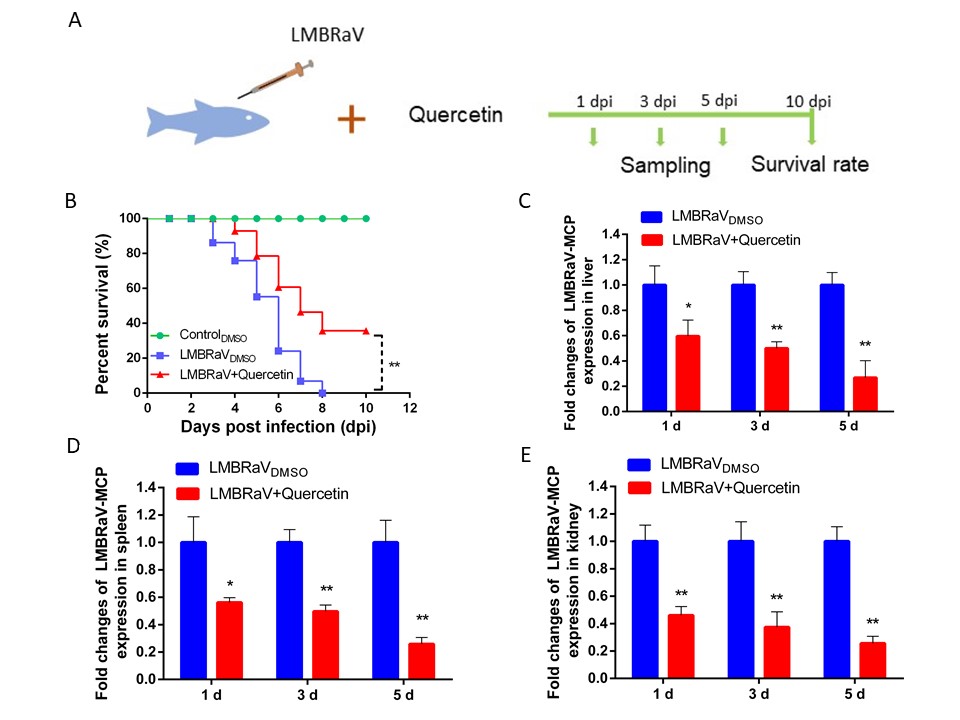
Protective effects of quercetin on LMBRaV-infected fish
In our in vivo experiments, we evaluated the potential toxicity of quercetin by observing clinical manifestations. Specifically, quercetin doses above 50 mg/kg resulted in slower swimming speed and reduced food intake for largemouth bass.
The results of quercetin’s protective effect on largemouth bass infected with LMBRaV are shown in Figure 5. The largemouth bass infected with LMBRaV exhibited a cumulative mortality rate of 100% after 8 days post-infection. Conversely, the quercetin-treated groups maintained an 80% survival rate at 5 days after infection, with a final survival rate of 40% at the conclusion of the experiment. The results of RT-qPCR assay showed that the relative expression levels of LMBRaV MCP gen in the liver, spleen and kidney of largemouth bass in the quercetin feeding group decreased significantly at 1, 3 and 5 d, suggesting that quercetin has a good therapeutic effect on LMBRaV infection in largemouth bass.
_flow_chart_of.jpeg)
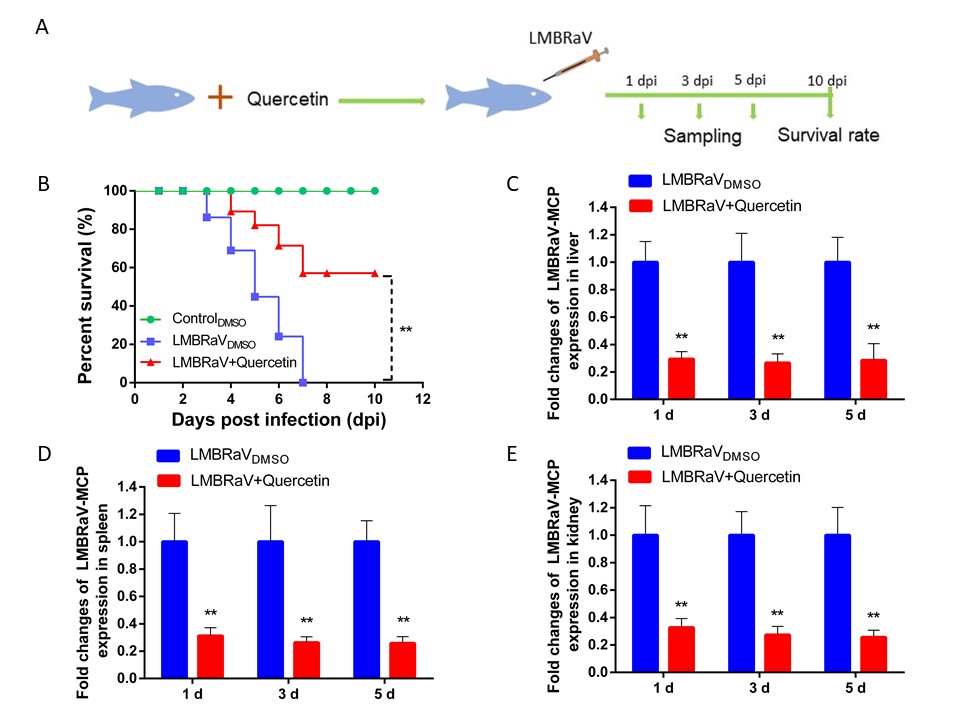
The preventive efficacy of quercetin against LMBRaV infection
Upon observing the effective treatment of quercetin on LMBRaV-infected largemouth bass, we delved into the inhibitory impact of quercetin pretreatment on LMBRaV infection.
The cumulative mortality rate of largemouth bass in the LMBRaV-infected group was 100% at 7 days after infection. Specifically, treatment with quercetin improved the survival rate of largemouth bass by 57.7%. Meanwhile, the relative expression of LMBRaV MCP gen in the liver, spleen and kidney of largemouth bass in the quercetin treatment group decreased significantly at 1, 3 and 5 d.
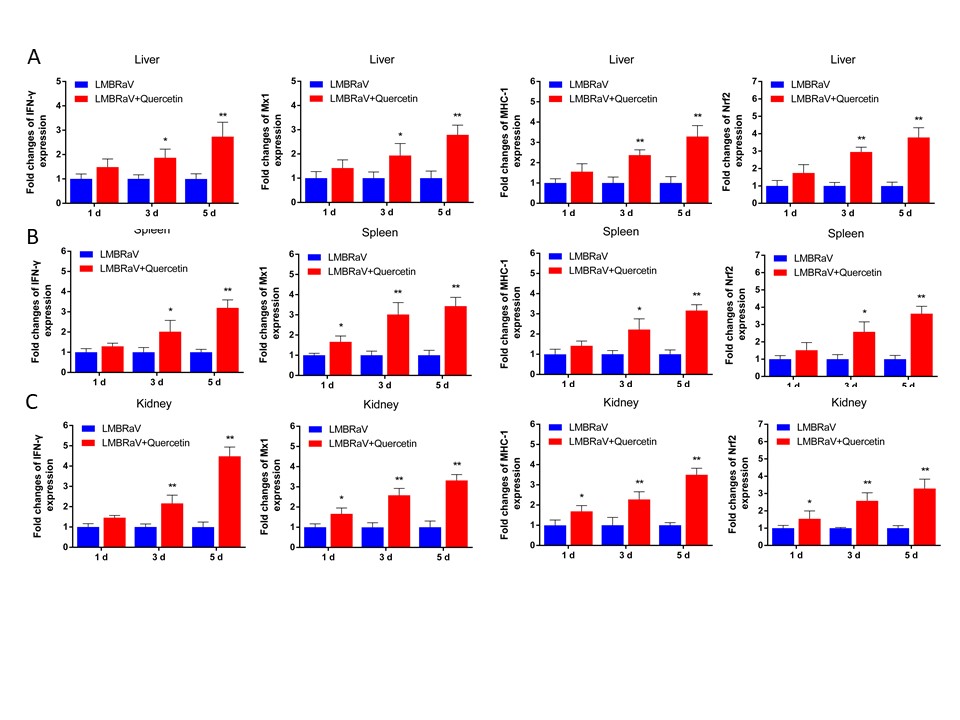
Expression of immune related genes
To investigate the potential antiviral mechanism of quercetin in largemouth bass, we analyzed the expression of immune related genes. Our findings revealed a significant up-regulation in the expression levels of key genes, such as IFN-γ, Mx1, MHC-1, and nuclear factor erythroid 2-related factor 2 (Nrf2), in LMBRaV-quercetin group (Figure 7). The up-regulation of Mx1, MHC-1, and Nrf2 expression levels was observed on days 3 and 5 following treatment with LMBRaV-quercetin. Particularly noteworthy was the significant increase in expression levels on day 5, with factors of 2.8, 3.3, and 3.7 observed in the liver. Similar trends were also observed in the spleen and kidney.
_flow_cha.jpeg)

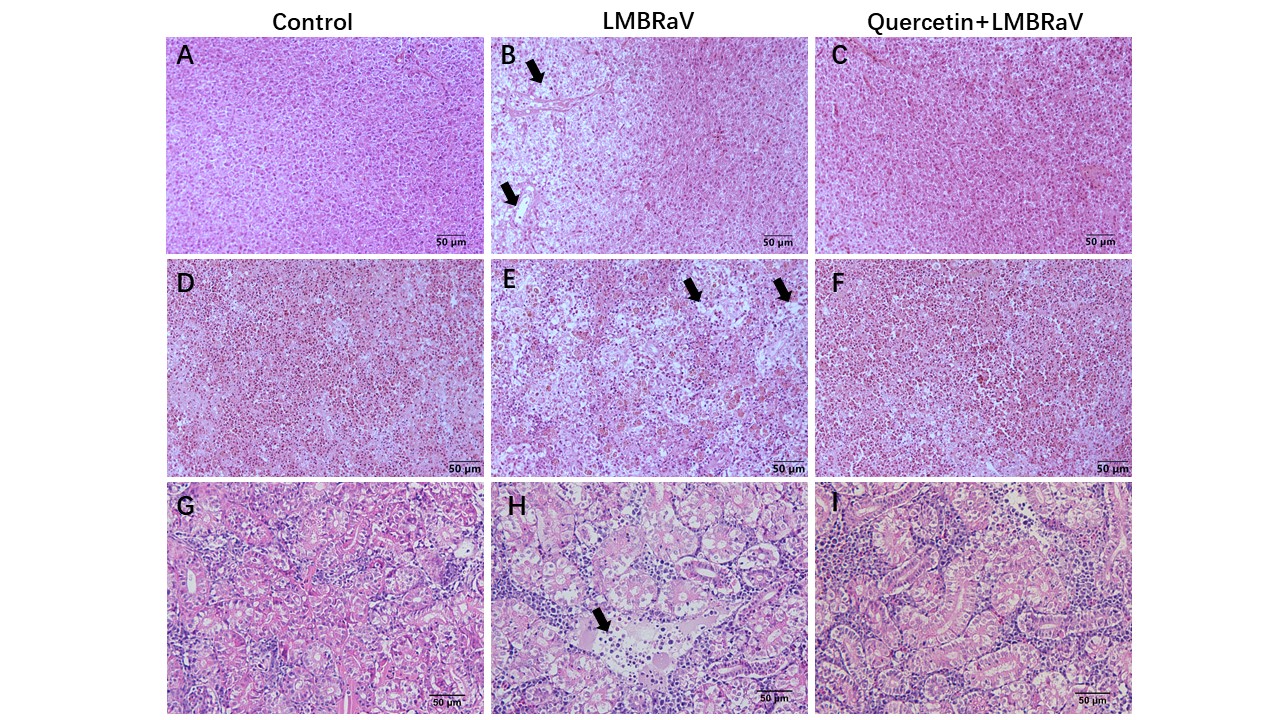
Histopathological observation
The effects of quercetin on histopathological alterations in immune organs of largemouth bass infected with LMBRaV were illustrated in Figure 8. In the group infected with LMBRaV, liver cells displayed pronounced necrosis, spleens showed marked vacuolization with blurred demarcation between red and white areas, and kidney tissues exhibited severe necrosis. Conversely, in the group treated with quercetin, no significant variance was observed compared to the control group, suggesting the potential of quercetin to alleviate LMBRaV-induced pathological harm to immune organs.

Discussion
M. salmoides has been widely utilized in global commercial aquaculture.20 The largemouth bass aquaculture industry faces significant challenges, particularly from viral diseases such as LMBRaV, necessitating urgent attention. Currently, there is a lack of effective drugs to control LMBRaV infection in largemouth bass.8,19 Therefore, developing a effective method to resist LMBRaV is crucial. Natural plants and their bioactive compounds have been extensively studied for their potential in treating viral diseases.9,21 Previous research has demonstrated the promise of identifying anti-LMBRaV compounds from natural sources.9,19 This study examined the anti-LMBRaV activity of nine natural products. The findings indicate that quercetin possesses previously unreported anti-LMBRaV properties. Quercetin showed significantly inhibited LMBRaV MCP gene expression in EPC cells, maintained the normal morphology of EPC cells, and exhibited concentration-dependent antiviral effects. Notably, at a concentration of 20 mg/L, quercetin demonstrated the highest inhibition rate against LMBRaV, reaching 93.6%. Quercetin is an environmentally friendly natural flavonoid compound that is widely found in apples, onions, tea, and red wine.22 According to previous reports, quercetin can inhibit the growth of Aeromonas hydrophila,23 suppress Grouper Iridovirus infection,24 and has inhibitory effects on White Spot Syndrome Virus in crayfish25 and genotype II Grass Carp Reovirus in Gobiocypris rarus.26
Apoptosis are frequently observed consequences of viral infections, involves a series of intricate molecular mechanisms and signaling pathways, yet targeted antiviral medications have been shown to alleviate these effects.27 Studies have demonstrated that LMBRaV infection induced typical apoptosis in EPC cells through intrinsic and extrinsic pathways.28 The present investigation reveals that quercetin provises significant protection against the morphological and nuclear fragmentation induced by LMBRaV infection in EPC cells, thereby blocking host cell cytopathic effects and cell death induced by LMBRaV.
Virus infection begins by attaching to the surface of a host cell, penetrating the cell, releasing genetic material, and replicating their genome.7,29,30 Antiviral medications predominantly impede or eliminate viruses through three principal mechanisms31,32: direct interaction resulting in viral destruction, interference with viral attachment to host cells, or inhibition of viral replication. Additionally, antiviral agents can attenuate host cell damage during viral infection by modulating host cell adherence and endocytosis.17 Several natural small molecules have been found to inhibit viral infections by blocking the entry of viruses into host cells.19,33 Tetrandrine has been shown to decrease SVCV infection by interfering with viral internalization.17 Quercetin inhibits Hsp70-mediated obstruction of Bovine Viral Diarrhea Virus infection and replication in the initial stages of infection.34 Small-molecule antiviral coumarin derivatives sourced from diverse plant species have demonstrated efficacy in disrupting multiple stages of the viral life cycle, including binding, internalization, and post-entry transport.31,35 This study demonstrates the significant reduction of LMBRaV infectivity by quercetin, as well as its inhibitory effects on infection through imposing LMBRaV entry into host cells. Additionally, virus-quercetin co-incubation experiments demonstrated quercetin’s direct virucidal effects. These results also reveal that quercetin inhibits LMBRaV through multiple pathways.
In vivo experiments showed that the preventive protection rate of quercetin against LMBRaV infection in largemouth bass was 57.7% and the therapeutic protection rate was 40%. This results indicated that quercetin had better preventive effect than therapeutic effect on LMBRaV infection in largemouth bass. Additionally, quercetin demonstrated a significant reduction in viral load. Previous studies have reported that saikosaponin C can increase the survival rate of largemouth bass infected with LMBRaV by 83.3%.10 Under laboratory artificial infection conditions, the anti-LMBRaV effect of saikosaponin C is better than that of quercetin. However, its effectiveness in natural aquaculture settings still requires further verification. Drugs may exert antiviral effects by interfering with viral replication or activating host immunity.36 Immunostimulants have the ability to enhance both nonspecific and specific defense mechanisms in fish, thereby enhancing their immune competence and resistance to diseases.17 Emerging evidence indicates that natural small molecules have the potential to enhance various immune responses, including activating the immune pathway, promoting antibody production, and activating the lysosomal system.37,38 Prior research has demonstrated that artemisinin could against SVCV infection by upregulating immune-related genes such as type I IFN (IFN1), IFN-stimulated gene product 15 (ISG15), Mx1, and viperin.39 Studies have also shown that coumarin can inhibit pathogen infection by increasing the expression of antimicrobial peptide-related genes.40 In the present investigation, the administration of quercetin in largemouth bass resulted in a notable upregulation of immune-related genes, such as IFN-γ, MHC-1, and Mx1. Nrf2 is involved in activating a series of genes related to cell protection and antioxidant enzymes. In addition, Nrf2 plays a key role in the regulation of inflammatory response in the initial stage of viral infection.41 The present study found that quercetin upregulated the expression of Nrf2, indicating that quercetin may enhance the host’s immune response by promoting the expression of immune related genes. These results indicated that quercetin could be used as an immunostimulant to prevent and treat viral diseases in aquaculture. Histopathological analysis demonstrated that LMBRaV can induce significant tissue damage in immune organs of largemouth bass, while quercetin has the ability to mitigate this pathological damage caused by LMBRaV. It is worth noting that in natural aquaculture settings, factors such as the aquatic environment, temperature fluctuations, the types and quantities of pathogens, and human interventions can all affect the immune response of largemouth bass, thereby influencing the efficacy of antiviral drugs.
In conclusion, this study demonstrates the efficacy of quercetin in inhibiting LMBRaV replication both in vitro and in vivo. Moreover, quercetin maintains cellular morphology, reduces LMBRaV-induced nuclear damage, and inactivate the virus directly. In addition, quercetin enhanced the innate immune response of largemouth bass, indicating its promise as an immune stimulant to prevent disease outbreaks in largemouth bass farming.
Acknowledgments
This work was supported by the Science and Technology Innovation Yongjiang 2035 Key Research and Development Project of Ningbo (2024Z279), the National Key Research Development Program of China (2023YFD2400704), the National Natural Science Foundation of China (32202980), and the Central Public-interest Scientific Institution Basal Research Fund (2020TD44).
Authors’ Contribution
Conceptualization: Mingyang Xue (Equal), Yong Zhou (Equal). Data curation: Mingyang Xue (Equal), Yangyang Jiang (Equal), Zhenyu Huang (Equal). Investigation: Mingyang Xue (Equal), Nan Jiang (Equal). Methodology: Mingyang Xue (Equal), Yangyang Jiang (Equal), Yong Zhou (Equal). Project administration: Mingyang Xue (Equal), Zhenyu Huang (Equal), Yong Zhou (Equal). Writing – original draft: Mingyang Xue (Equal), Yangyang Jiang (Equal). Formal Analysis: Yangyang Jiang (Equal), Chen Xu (Equal), Yan Meng (Equal). Resources: Chen Xu (Equal), Zhenyu Huang (Equal), Nan Jiang (Equal). Validation: Chen Xu (Equal), Nan Jiang (Equal), Yiqun Li (Equal). Visualization: Chen Xu (Equal), Zhenyu Huang (Equal), Yan Meng (Equal). Software: Nan Jiang (Equal), Yiqun Li (Equal), Yan Meng (Equal). Funding acquisition: Yiqun Li (Equal), Yuding Fan (Equal), Yong Zhou (Equal). Supervision: Yiqun Li (Equal), Yuding Fan (Equal), Yong Zhou (Equal). Writing – review & editing: Yuding Fan (Equal), Yong Zhou (Equal).
Competing of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical Conduct Approval
All animal experiments were conducted in compliance with the Ethical Inspection for Animal Experimentation (Ethical protocol code: YFI2024-zhouyong-0125).
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.