


Introduction
As an aglycone metabolite of hesperidin, hesperetin (3′,5,7,-trihydroxy-4′-methoxyflavanone) is the most abundant flavonoid glycoside isolated from citrus fruits.1 Among them, rhamnose can also be removed by glycosyl hydrolase to form hesperetin.2 Hesperetin has higher bioavailability and has shown potential health-promoting properties on cancer, cardiovascular diseases, carbohydrate dysregulation, bone health, cardioprotective and neuroprotective function, and other pathologies.1,3 However, further well-designed experimental trials are needed to address other effects of hesperetin observed in preclinical trials, as well as to a more in-depth understanding of its safety profile.
Siniperca chuatsi rhabdovirus (SCRV) is one of the strains of fish rhabdoviruses, which are epidemic pathogens that harm fish aquaculture and cause great economic losses.4 However, there are currently no specific drugs to treat these diseases. Considering their enormous infectivity and damage to various fish species, a large number of studies have been reported on the molecular biology of rhabdoviruses and their interactions with hosts,5–7 which provided new information for understanding viral infection mechanism and developing potential antiviral techniques against fish rhabdoviruses. For SCRV virus, previous research showed that its infection could activate apoptosis and autophagy,8 interplay of which serves an important role in the virus replication and antiviral immune of hosts.6 Transcriptome analysis of immune response against SCRV in infected fish tissues and microRNAs profiles of SCRV-infected cells were also investigated.9,10 These studies provided further profound insight into the antiviral immune response and pave new ways for the development of effective strategies against SCRV infection.
In this study we explored the antiviral activity of hesperetin against SCRV in vitro. Our current study demonstrated that hesperetin could apparently delay the cytopathic effects induced by SCRV in cell monolayers and inhibit the replication of progeny virus. Further mechanical research suggested that hesperetin could block the intrusion of SCRV into cells after infection. Hesperetin also led to a decrease in apoptotic cells and protected the cells from SCRV infection. This study undoubtedly provides a good preliminary foundation for the research and development of antiviral technology based on hesperetin against fish rhabdoviruses.
Materials and methods
Cells and SCRV maintenance
EPC (epithelioma papulosum cyprinid) cells used in this study were cultured in M199 medium (Haiken, China) containing 10% fetal bovine serum (FBS, Sijiqing, Hangzhou, China) and maintained at 25°C in an incubator. The SCRV virus was stored in freezer at -80°C.
MTT assay
EPC cells were seeded in sealed culture dishes for growth overnight, resulting in about 75% to 85% confluence of the culture dish. 4 ml of different concentrations of hesperetin solution (0, 150, 300, 400 μM) was added to each dish for 24 and 48 h, respectively. The toxicity of the hesperetin to the cells was detected by MTT assay according to standard procedures.11 In addition, the morphology changes in treated cells were observed by inverted microscope at different time points.
CPE detection and time-of-addition assay
After growing to about 75% to 85% confluence of the culture dish, EPC cells were infected with SCRV (350 TCID50) for 2 h, or incubated with 4ml of different concentrations of hesperetin (0, 150, 300, 400 μM) containing SCRV (350 TCID50) solution prepared with cell culture medium (containing 5% FBS), respectively. CPEs of cell monolayers were observed by inverted microscope and photographed at different time periods.
For time-of-addition assay, EPC cells were treated with below three different infection procedures, including neutralization group (co-treatment: mixture with hesperetin and SCRV), inhibition group (post-treatment: firstly, infected with SCRV, then added hesperetin), and blocking group (pre-treatment: firstly incubated with hesperetin, then infected with SCRV). After 48 h, the cell supernatant and cell lysate of each group were taken to further detect the virus titer, respectively. The TCID50 values of the virus solutions were calculated by standard Reed-Muench method.11
Western blotting
The treated EPC cells were collected for protein extraction. After the determination of protein concentration by the Coomassie brilliant blue method, equal amounts of protein samples were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to nitrocellulose membranes. After blocking with 5% skimmed milk powder, the membranes were incubated with different primary antibodies overnight at 4 °C, and further incubated with the corresponding horseradish peroxidase (HRP)-conjugated secondary antibodies at room temperature for 1 h, respectively. After washing three times with TBST solution, the membranes were detected with the Odyssey CLX infrared imaging system (Li-COR Biosciences, Lincoln, Nebraska USA), and the fluorescence intensities of the bands were quantified by ImageJ software.
qRT-PCR
Total RNA samples were extracted from treated EPC cells using the Animal Tissue Total RNA Extraction Kit (Prime Biotech), and then reverse-transcribed into cDNA according to the procedures of the Transcription Factor Kit (Yeasen Biotechnology Co., Ltd, Shanghai, China). qRT-PCR analysis was performed with the SYBR Green kit (Vazyme Biotech Co., Ltd. Nanjing, China) using 18s rRNA as an internal reference gene. The primers used in this study were as follows: SCRV N-F: ATCCATCAGATCACAGAACGC, SCRV N-R: TCCCAGCCATTCTCCTCAGTCC; 18s-F: CATTCGTATTGTGCCGCTAGA,18s-R: CAAATGCTTTCGCTTTGGGTC. The relative mRNA values of each group were calculated by 2-ΔΔCt method. The experiment was repeated three times in parallel.
Hoechst 33342 staining
To detect the nuclei changes in SCRV-infected EPC cells, after incubation with hesperetin and SCRV, the cells were stained with a Hoechst 33342 solution in the dark for 15 min. The nuclei of each group of cells were observed under a Zeiss fluorescence microscope.
Flow cytometry analysis
For apoptotic rate detection, the treated cells were collected and stained with Annexin V-FITC (5 μL) and PI (10 μL) solution successively, protected from light for 20 min, and then immediately detected by flow cytometry (BD FACSCanto™II Flow Cytometry, USA). The data were finally analyzed by Flowjo software.
Caspase activity detection
Enzyme activities of caspases (including caspase-3, caspase-8 and caspase-9) were detected in EPC cells treated with hesperetin and SCRV. The assay was carried out according to the protocol of the Enzymatic Activity kit Reaction System (Beyotime Biotechnology, Shanghai, China) and analyzed at a wavelength of 405 nm with the microplate reader (Bio-Tek Epoch, USA). The enzyme activities were calculated by the generated absorbances and the comparison standard curve.
Mitochondrial membrane potential (MMP) assay
After acting on SCRV-infected EPC cells with hesperetin for 24 h, cells were incubated with JC-1 staining (Beyotime Biotechnology, Shanghai, China). After washing twice with pre-cooled JC-1 staining buffer, the fluorescence signals of the cells were detected with a Zeiss fluorescence microscope.
ROS measurement
After 24 h of treatment with hesperetin in SCRV-infected EPC cells, the cells were washed with PBS solution and followed by DCFH-DA staining to detect changes in ROS levels. The fluorescence signals in the cells were observed and photographed with a laser confocal microscope (Olympus Fluorescence Vision FV3000, Japan).
Statistical analysis
Data are presented as mean ± standard error, and all experiments were repeated at least three times. All statistical analyses were performed with IBM SPSS Statistics 25 software. p < 0.05 was considered statistically significant (*p < 0.05; **p < 0.01, and ***p < 0.001).
Results
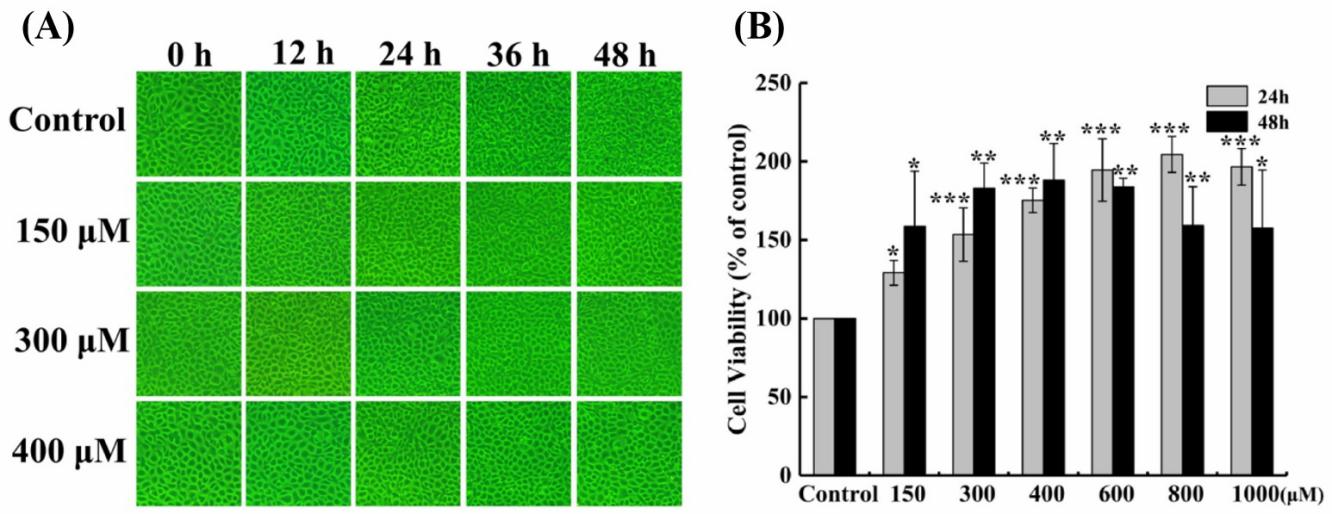
There’s no toxic effect of hesperetin to EPC cells
In order to explore the antiviral effect of hesperetin against fish rhabdovirus SCRV, we first detected whether it has a toxic effect on the growth of EPC cells or not. The current study showed that after treatment, the cell monolayer still remained very complete and did not change significantly with the increase of the concentration of hesperetin at different time periods (Figure 1A), and there’s no apparent toxic impact on the growth of EPC cells. Interestingly, the MTT assay suggested that the survival rate of EPC cells increased with the higher concentration of hesperetin (Figure 1B), indicating that hesperetin might have a promoting effect on the growth of normal cells.
_morphological_c.jpeg)
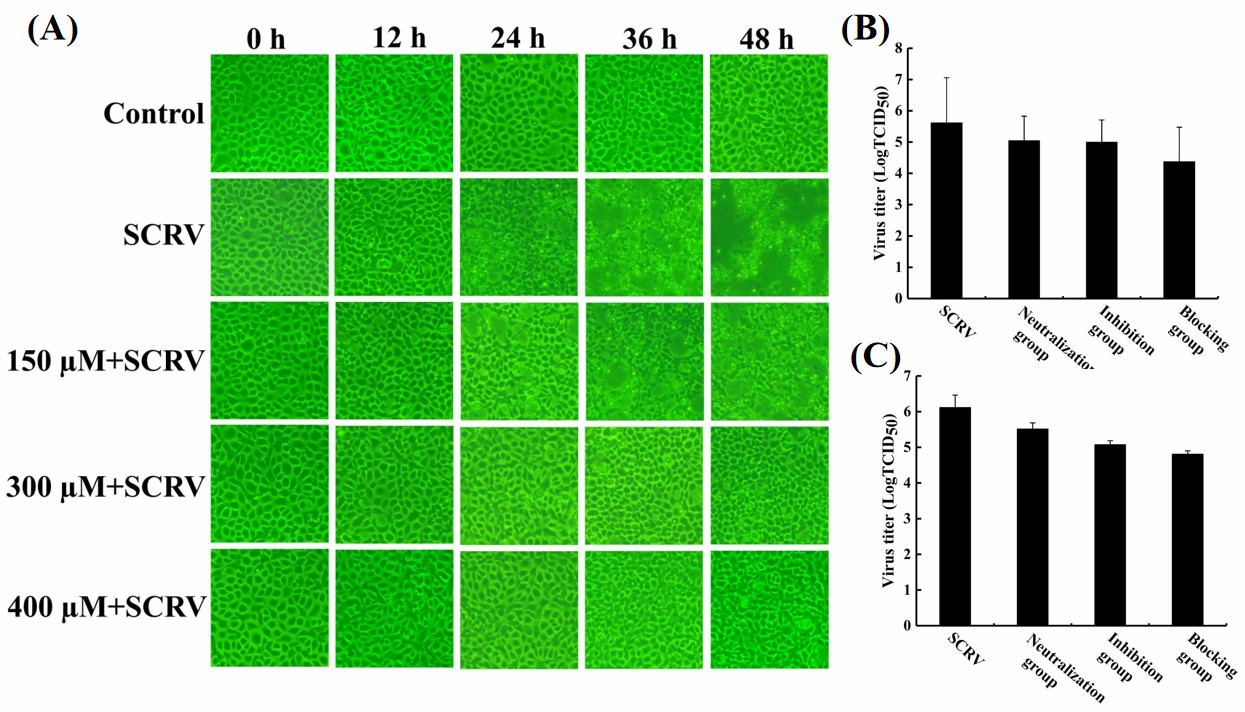
Hesperetin possesses a strong antiviral ability to SCRV
To investigate the antiviral ability of hesperetin to SCRV, different concentrations of hesperetin were applied to treat SCRV-infected EPC cells. Microscope observation found that the cell monolayer in the SCRV-only infection group showed more CPEs, and the cells shrank, brightened, and detached from the monolayer. While, with the increase of the concentration of hesperetin, the numbers of CPEs in the experimental groups were obviously less than that in the infection-positive group, with a time- and concentration-dependent manner (Figure 2A), indicating that hesperetin possessed strong anti-SCRV effect on EPC cells.
_microscopy_observation_of_scrv-.jpeg)
Besides, a time-of-addition experiment was explored to identify the antiviral mode of hesperetin against SCRV infection. After treatment with three different procedures (neutralization group, inhibition group, and blocking group), the virus titers in the cell supernatants (Figure 2B) and lysates (Figure 2C) at 48 h post-infection were tested. Compared with the SCRV-only infection group, the virus titers in the groups treated with hesperetin displayed a downward trend (Fig. 2B; 2C), and there’s no significant difference between the titer of supernatant and cell lysate in the same treatment group. Among the 4 groups, TCID50 in supernatant or lysate samples of the blocking group exhibited the lowest value, which suggested that hesperetin might execute its antiviral function by blocking SCRV attachment or entry into the cells.
Hesperetin inhibited expression of SCRV nucleoprotein
In order to explore the antiviral mechanism of hesperetin against SCRV, we detected the transcription and expression of the SCRV nucleoprotein gene (N gene), which plays a key role in rhabdovirus assembly and replication in infected cells. qRT-PCR analysis showed that under all four treatment conditions (SCRV-only infected group, neutralization group, inhibition group, and blocking group), the transcriptional level of SCRV N gene in the blocking group was lowest (Figure 3A). Furthermore, western blot and its statistical analysis indicated that N protein also exhibited the lowest expression manner in SCRV-infected EPC cells by pre-exposure of hesperetin (Figure 3B), which was consistent with qRT-PCR results and the morphological observation described above.
_qrt-pcr_analysis_of_scrv_*n*_gene_w.jpeg)
Hesperetin reduced apoptosis in SCRV-infected EPC cells
A previous study found that SCRV infection could induce cell apoptosis.11 In this study, Hoechst 33342 staining analysis confirmed the existence of typical apoptotic features such as partial rupture and crescent shape of the nucleus (Figure 4A), while pre-exposure of hesperetin could significantly reduce the apoptotic nuclei in the blocking group, compared to the only-SCRV infected group (Figure 4A), which indicated that hesperetin was able to decrease apoptosis caused by SCRV in EPC cells.
_hoechst_33342_staining_of_cell.jpeg)
In addition, the cell apoptosis rates in the blank control group, hesperetin-treated group, SCRV-infected group, and the blocking group, were counted by flow cytometry, respectively. The results showed that the percentage of apoptosis in the SCRV-alone infected group came to 18.84% (including early and late apoptotic cells) at 24 h, while the hesperetin could retard the cell apoptosis induced by SCRV to some extent, and the cell apoptosis rates were 11.78% (Figure 4B). This also reaffirmed that hesperetin could protect EPC cells from apoptotic cell death triggered by SCRV infection.
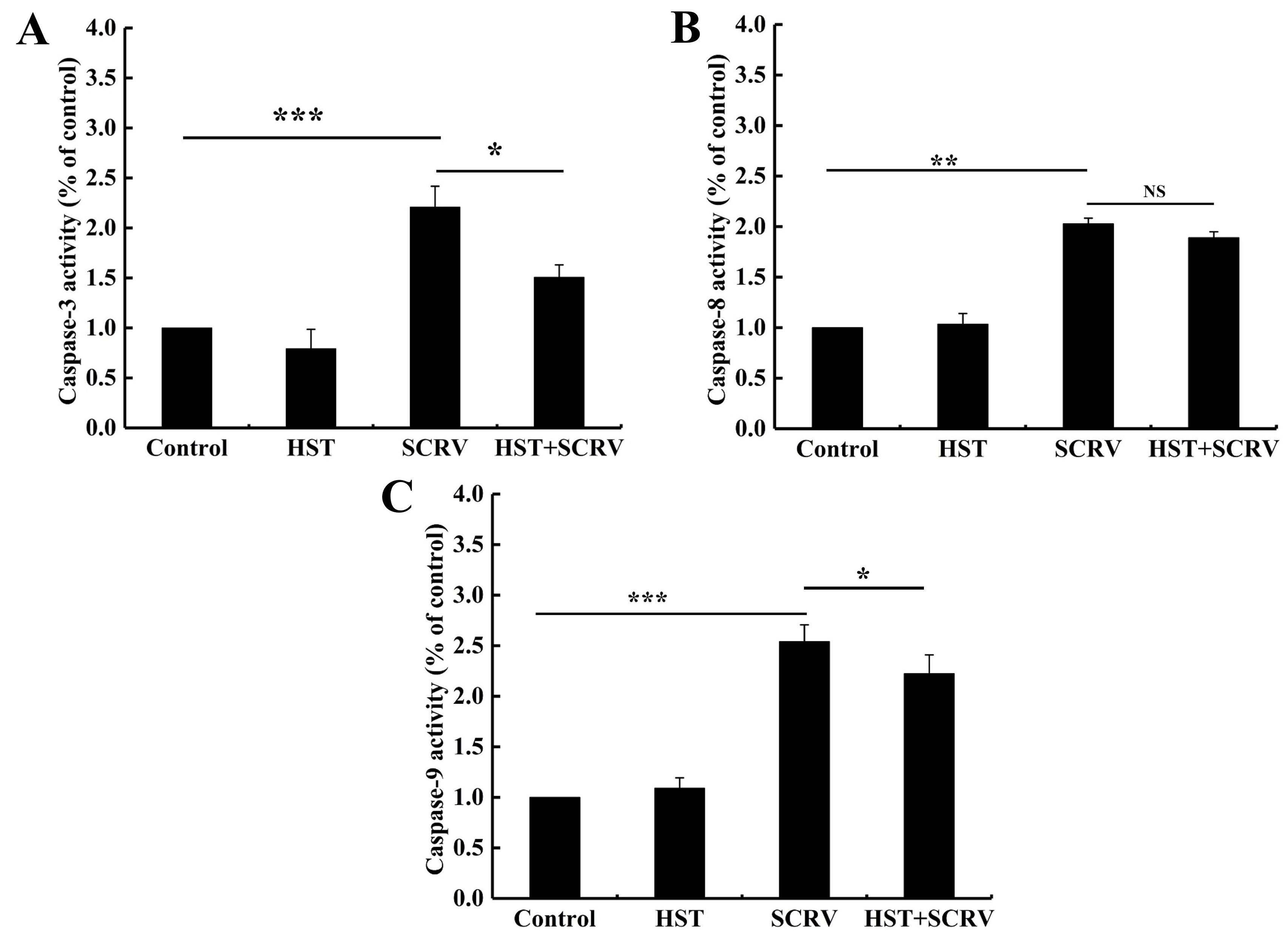
Hesperetin reduced caspase-3 and caspase-9 activity in SCRV-infected cells
The cystatin family of proteases is closely associated with apoptosis. The changes in enzymatic activities of the main executive caspases (caspase-3, caspase-8, and caspase-9) were examined in different test groups. The results showed that, after 24 h of post-infection, the caspase-3, caspase-8, and caspase-9 activity increased 2.5-fold, 2.1-fold, and 2.7-fold in the SCRV-infected group compared to the blank control group, respectively (Figure 5). Among them, hesperetin could significantly decrease the activity of caspase-3 and caspase-9 in the blocking group compared to the SCRV-only infected group, while there’s no statistical difference in caspase-8 activity between the two group cells (Figure 5). Thus, the current study suggested that hesperetin might block the intrinsic mitochondrial apoptosis pathway activated by SCRV infection in EPC cells.

Hesperetin inhibited mitochondrial apoptosis in SCRV-infected cells
To further confirm the inhibitory role of hesperetin in apoptosis induced by SCRV infection, we used DCFH-DA and JC-1 probe to detect changes in ROS and MMP of SCRV-infected EPC cells, respectively. After DCFH-DA staining, the number of green fluorescent signals significantly increased in SCRV-infected cells, while the number of positive signals decreased apparently in the blocking group (hesperetin + SCRV) (Figure 6A), which suggested that hesperetin pretreatment could disturb the SCRV-induced ROS rise in EPC cells.
_detection_of_ros_changes_in_epc_cells_with_different_exposure_procedures._pretreatment.jpeg)
Additionally, the JC-1 staining analysis demonstrated that the green fluorescence signals (low MMP) were significantly enhanced in the SCRV-infected cells, which suggested the activation of cellular mitochondrial intrinsic apoptosis (generally, JC-1 could produce red light in the matrix with high MMP, while the green light when the potential is low). However, pretreatment of hesperetin could efficiently increase red signals (high MMP) and reduce the generation of green signals activated by SCRV (Figure 6B). These data indicated that hesperetin could inhibit mitochondrial apoptosis in SCRV-infected EPC cells.
Discussion
Flavonoids are a diversified group of natural substances that were discovered to provide a variety of health benefits in human beings, including hepatoprotective properties, anticancer, anti-inflammatory, and antiviral activities.12,13 Isolation, identification, and biological role of flavonoids, as well as their uses on health advantages, are all major topics in research and development currently. Flavonoids have been studied to fight against a variety of DNA and RNA viruses. For example, quercetin, apigenin, and proanthocyanidins have been investigated for their antiviral effects on influenza virus type 3 and herpes simplex virus.14 In view of the arrangement, replacement of functional groups, and total number of hydroxyl groups around flavonoid’s nucleus structure affecting their biological activity, metabolism, and bioavailability, more studies on the pharmaceutical properties of different flavonoids need to be investigated.
In this study, hesperetin, the natural flavanone compound, was first used to discover its effects against the fish rhabdovirus SCRV, one major threat to hinder the development of aquaculture, causing severe hemorrhagic sepsis in freshwater and marine fish and several economic losses.15,16 Through a combination of morphological observation, virus titer assay, qRT-PCR and western blot analysis, hesperetin was found to inhibit CPE occurrence, lower the progeny virus titer and decrease the expression of SCRV N protein. Additionally, in view of the complex process including virus adsorption, penetration, replication of the parental virus, and release of progeny virus in the viral replication cycle, time-of-addition assay was conducted, and our study suggested that pretreatment of hesperetin could efficiently block the SCRV attachment or entry and therefore interfere with the replication of progeny viruses in EPC cells.
Apoptosis accounts for a large proportion of several types of cell death when rhabdoviruses infect host cells.17 Zhou et al. (2021)8 also proved that SCRV infection can cause apoptosis of EPC cells. In the present study, by Hoechst 33342 staining, Annexin V/PI double staining and detection of activity of apoptosis-related enzymes, hesperetin was found to reduce the endogenous apoptosis rate of SCRV-infected EPC cells and exhibit antiviral activity by preserving mitochondrial integrity and reducing ROS in SCRV-infected EPC cells. Endogenous apoptosis can lead to mitochondrial dysfunction, and changes in MMP and ROS levels are key features. Hence, the present JC-1 and DCFH-DA fluorescence probe staining analysis also confirmed that hesperetin could reduce mitochondrial damage and prevent endogenous apoptosis caused by SCRV infection in EPC cells. These results could help to explore profoundly the antiviral mechanism of hesperetin to fish rhabdoviruses in the future.
In fact, previous studies have confirmed that hesperetin could inhibit replication of a variety of viruses, such as herpes simplex virus type 1, poliovirus type 1, parainfluenza virus type 3, influenza virus, Kaposi’s sarcoma-associated herpesvirus, severe acute respiratory syndrome coronavirus 2 and Dengue virus,14,18–21 and affect multiple viral infection processes and regulate intracellular signal pathways involved in cell proliferation and responses to environmental changes.20 In aquatic virology, hesperetin was also found to protect crayfish against white spot syndrome virus (WSSV) infection, the important invertebrate pathogen in aquaculture.22 It can regulate the innate immunity of crayfish and delay and reduce mortality after the WSSV challenge. Recently, Verma et al. (2024)23 reported the antiviral activity of hesperetin against multiple poxviruses, including buffalopox virus, vaccinia virus, and lumpy skin disease virus, providing some insights into its potential mechanisms of action. There is no doubt that our current study provides new data and evidence on the antiviral spectrum of hesperetin.
However, we should note that SCRV replication cycle is a complicated process involving multiple factors in infected cells.24 Although some new molecules against SCRV infection have been reported,11 more and more antiviral drugs or techniques need to be explored in the future. Obviously, current findings on the dual role of hesperetin in both directly suppressing SCRV replication and modulating the cell mitochondrial apoptosis, offer a promising candidate molecule and improve the management of SCRV infection, ensuring healthier fish populations and greater economic stability.
Acknowledgments
This research was supported by Natural Science Foundation of Henan (232300420018) and the High-level talents Fund from Henan University of Technology (2023BS001).
Authors’ Contribution
Conceptualization: Yu-Zhen Ma (Equal). Formal Analysis: Yu-Zhen Ma (Equal), Shuang-Shuang Sun (Equal). Investigation: Yu-Zhen Ma (Equal), Shuang-Shuang Sun (Equal). Writing – original draft: Yu-Zhen Ma (Equal), Shuang-Shuang Sun (Equal). Writing – review & editing: Shi-Wei Ma (Equal). Funding acquisition: Shi-Wei Ma (Equal). Supervision: Shi-Wei Ma (Equal).
Competing of Interest – COPE
The authors declare that they have no financial/commercial conflicts of interest concerning this article.
Ethical Conduct Approval – IACUC
This study does not involve in vivo experiments on animals or plants. All analyses were based on previously published studies. Thus, no ethical approval and patient consent are required.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All data generated or used during the study appear in the submitted manuscript.