

1. Introduction
Culter alburnus is an economically valuable freshwater fish found in China.1,2 Megalobrama terminalis is an important fish species have delicious flesh.3 C. alburnus (♀) × M. terminalis (♂) (CM) is a new national variety formed by interspecific hybridization between C. alburnus and M. terminalis , with fast growth, low demand for feed protein and high economic value.4 Liu et al.3 determined the complete mitochondrial DNA sequence of CM. The results showed that sequence alignment between the mitochondrial genomes of the hybrid and its female parent showed that a total of 35 mutation sites were identified in 14 genes or regions. However, there is relatively little research on other aspects of CM. On the other hand, in recent years, a multivariate analysis method combining measurable traits and framework structure parameters has achieved good results in distinguishing fish morphological differences.5 Principal component and discriminant analysis methods are widely used. Cluster analysis intuitively analyzes the differences and connections between the classification objects. Principal component analysis can summarize the morphological differences between different groups, and using discriminant functions and corresponding measurement indicators, any undetermined sample can be classified into one of the groups.6,7 Therefore, it was necessary to conduct comparative studies on the appearance characteristics of CM and C. alburnus to provide an important biological basis for germplasm identification, targeted breeding, and improved variety breeding of CM and C. alburnus, and to accumulate data for genetic research on distant hybridization of fish.
During the production process of fish farming, abnormal weather, sudden changes in water quality, and prolonged transportation times can cause a decrease in the DO value of the water body, causing fish to face low-oxygen conditions.8,9 Fish maintain normal life activities by altering the activities of related enzymes and other hypoxic stress responses in hypoxic environments.10 However, excessive stress can cause damage to tissues and organs as well as an imbalance in the homeostasis of the fish’s internal environment and immune system, thereby affecting growth and survival.11 Owing to the sensitivity of fish belonging to the subfamily Culter in hypoxic environments, researchers have researched the survival status of fish belonging to the subfamily Culter under hypoxic conditions. Cheng et al.12 studied the effects of different levels of DO on physiological indicators, antioxidant enzyme activity, and liver histology of C. alburnus and “All-female No.1” C. alburnus and speculated that the tolerance of “All-female No.1” C. alburnus to hypoxic environments might be slightly higher than that of C. alburnus. Zhu et al.13 measured the oxygen consumption rate and suffocation point of C. alburnus fry at different temperatures, and their results showed that the oxygen consumption and oxygen consumption rate of C. alburnus fry of the same specifications increased with increasing water temperature, showing a positive correlation. The suffocation point increases slightly with increasing water temperature. Li et al.14 found that the suffocation point of Ancerythricuter nigrocauda slightly increased with the increase in water temperature and fish size, besides, it was found that in water temperatures ranging from 14 ℃ to 32 ℃, the oxygen consumption rate of Ancerythricuter nigrocauda showed significant periodic changes, which could guide artificial breeding based on their activity patterns. However, previous studies mainly focused on the changes in oxygen consumption rate and asphyxiation point of Culter subfamily fishes in the hypoxic environment, and there was no relevant study on the newly cultivated national species of CM. Thus, it was necessary to explore CM. In conclusion, this study analyzes the impact of DO level on C. alburnus and CM from the physiological ecology and biochemistry perspective. Through the determination of floating point, coma point, and asphyxiation point, and the determination of changes in Superoxide dismutase (SOD), Glutathione (GSH), and Malondialdehyde (MDA) activity of oxidative stress-related enzymes, exploring the physiological response and tolerance of C. alburnus and CM in low DO environment to guide variety breeding and response to hypoxia and other emergencies in the breeding process. It also provided a basis for the optimal breeding and healthy breeding of new varieties such as CM.
2. Materials and methods
2.1. General background
The experiments were conducted at the Zhejiang Institute of Freshwater Fisheries in Huzhou, China. The CM and C. alburnus used in this study were all healthy specimens of the same age.
2.2. Experiment 1: Study on morphological characteristics of CM and C. alburnus
A total of 50 tails CM were randomly selected, with an average body length of 32.19±2.66 cm and an average body weight of 493.47±35.21 g. A total of 50 tails C. alburnus were randomly selected with an average body length of 30.91±2.71 cm and an average body weight of 419.51±42.29 g. Nine quantifiable traits were measured (total length, body length, head length, snout length, tail stalk length, tail stalk height, body height, body thickness, and intestinal length), and 12 countable traits (lateral line scale, lateral line upper scale, lateral line lower scale, dorsal fin spine number, dorsal fin spine number, pectoral fin spine number, pectoral fin spine number, abdominal fin spine number, anal fin spine number, anal fin spine number, and first-gill rake number). Countable traits were directly counted, and measurable traits were measured using a Vernier caliper with measurement parameters accurate to 0.01 cm. To eliminate the impact of different fish body sizes on quantifiable trait parameters, quantifiable trait parameters were converted into proportional traits (total length/body length, body length/height, body length/body thickness, body length/head length, head length/snout length, head length/tail stem length, tail stem length/tail stem height, and body length/intestine length) for correction.
2.3. Experiment 2: Study on hypoxia tolerance of CM and C. alburnus
A total of 60 tails CM were randomly selected, with an average body weight of 4.99±0.12 g, and a total of 60 tails C. alburnus were randomly selected with an average body weight of 5.06±0.15 g.
2.3.1. Determination of physiological and ecological indicators of CM and C. alburnus
A 15-L suction bottle was used as the breathing chamber, and the bottle was sealed. Each suction bottle was filled with 20 similarly-sized CM or C. alburnus experimental fish, with a water temperature of 25 ± 0.5 °C (initial DO value of 6.5 mg/L), each in parallel with three groups. The reaction of the experimental fish was carefully observed, and the DO value at the floating head of the first fish was used as the floating head point. The DO value of the first fish, when it lost balance and fell unconscious on its side, was the critical point of the coma. The DO value at which 50% of the fish became unconscious was considered the coma point. The DO value of the first fish to die was the critical point of suffocation. A DO value of 50% of the fish deaths was used as the suffocation point. The DO value of all dead fish was considered the point of death. Fish lying still at the bottom of the bottle and only breathing through their gills were considered unconscious. If the gills and tail stopped moving and there was no response when lightly touched with a glass rod, death was considered. During the experiment, without feeding or changing the water, the DO values of the floating head point, coma critical point, suffocation critical point, coma point, suffocation point, and death point of the CM or C. alburnus were measured.
2.3.2. Detection of SOD, GSH, and MDA activities under hypoxia stress of CM and C. alburnus
A 15-L suction bottle was used as the breathing chamber, and the bottle was sealed. Twenty similarly-sized CM or C. alburnus experimental fish were placed in each suction bottle. The inlet pipe of the suction filter bottle was connected to the upper end of the flowmeter, and the outlet of the water storage bucket was connected to the lower end of the flowmeter. Thirty fish with normal body color, healthy and lively, and similar specifications were placed in each suction filter bottle and three parallel CM or C. alburnus groups. Controlled the inlet flow rate to maintain the DO value in the suction bottle at 2.0 ± 0.5 mg/L, water temperature at 25 ± 0.5 °C, and hypoxia stress for 0, 6, 12, and 24 hours. Taken the liver of CM or C. alburnus, stored them in liquid nitrogen, and then transferred them to an -80 ℃ ultra-low temperature refrigerator for storage. The tissue homogenate was prepared by adding 9 mL of physiological saline to each gram of the sample tissue, ground, and transferred to a 2-mL centrifuge tube. They were centrifuged at 3000 r/min at 4 ℃ for 10 minutes. The supernatant was taken and divided into several batches before temporarily storing it in a 4 ℃ refrigerator for measuring SOD, GSH, and MDA activities.
2.4. Statistical analysis
All data were presented as mean ± SD. After arcsine transformation, the data obtained from each experiment were analyzed by analysis of variance (ANOVA) using Statistical Product and Service Solutions (SPSS) software (version 17.0; IBM, Armonk, NY, USA) to determine group differences. Tukey’s multiple comparison test was performed as a post hoc test. For all tests, statistical significance was set at P ≤ 0.05.
3. Results
3.1. Characteristics of CM and C. alburnus
3.1.1. Comparison of main morphological features of CM and C. alburnus
The body was long and flat in the CM, similar to that of C. alburnus, but its height and thickness increased significantly. The head of CM is smaller and blunter than that of C. alburnus, featuring a prominent protrusion at the back. Additionally, CM has a significantly lower head length-to-body length ratio. The mouth was slightly longer than the upper forehead with oblique fissures. The eyes were equal in size. The lateral scale was relatively straight and ran through the middle of the fish body, similar to C. alburnus. The abdominal ridge extends from the abdominal fin base to the anus. The back and upper parts of the body were greyish-brown, and the abdomen was silver-white, similar to C. alburnus. The scales on the sides of the body were relatively large, unlike those in C. alburnus. The scales were thin and soft, with a few light melanin spots distributed on the side of the body. Each fin was gray, with gray-black ends and edges. In a word, the main characteristics of CM that were different from C. alburnus in appearance were the increase of body height and body thickness, small blunt head, bulge at the back of the head, reduced ratio of head length to body length, and larger scales at the side of the body (Figure 1).
_and_*c._alburnus*_(right_side).png)
3.1.2. Comparative analysis of conventional quantifiable traits of CM and C. alburnus
The results showed that there were significant differences (P<0.05) in body length/body height, body length/body thickness, body length/tail handle length, tail handle length/tail handle height, body length/gut length between C. alburnus (♀) × M. terminalis (♂) and C. alburnus, however, there were no significant differences in the three personality indicators of total length/body length, body length/head length, head length/snout length (P>0.05). Among them, body length/height, body length/thickness, tail handle length/tail handle height, and body length/gut length of CM were significantly smaller than those of C. alburnus (P<0.05), while body length/tail handle length was significantly larger than that of C. alburnus (P<0.05) (Table 1).
3.1.3. Comparative analysis of conventional countable traits of CM and C. alburnus
The results showed that the countable traits of CM and C. alburnus were significantly different (P<0.05) in the number of lateral scales, upper lateral scales, lower lateral scales, and first gill harrow. The number of lateral scales, the number of upper lateral scales, and the number of first-gill harrow of CM were significantly lower than that of C. alburnus (P<0.05); however, the number of lower lateral scales was significantly higher than that of C. alburnus (P<0.05). There was a slight difference in the number of pectoral and anal fins, whereas the other indicators were consistent (Table 2).
3.2. Hypoxic tolerance of CM and C. alburnus
3.2.1. Detection of floating point, coma point, suffocation point, etc., of CM and C. alburnus at 25 ℃
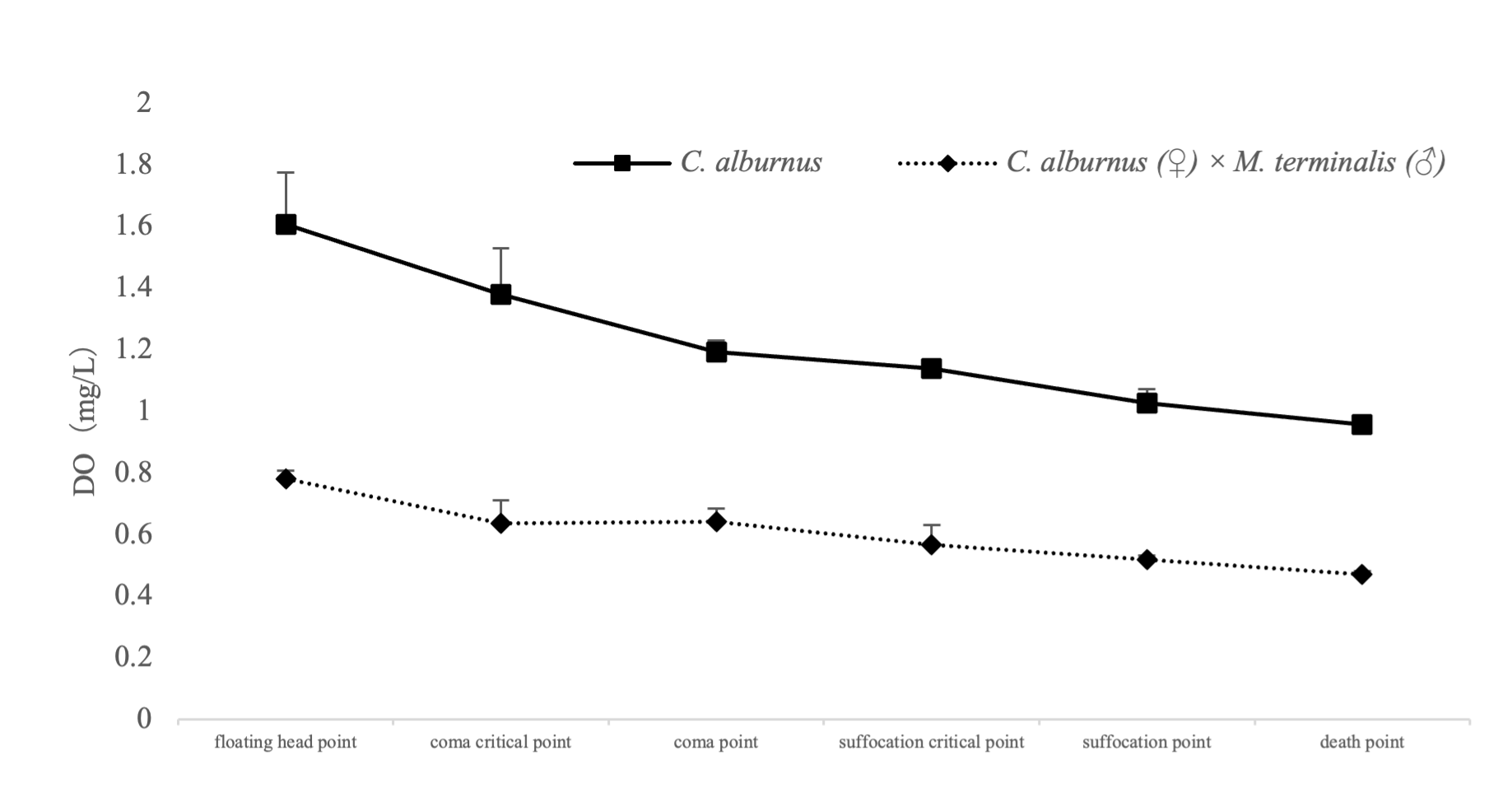
The results showed that the DO values of floating head point, coma critical point, coma critical point, asphyxia critical point, asphyxia critical point, and death point of CM were 1.333, 1.143, 0.997, 0.927, 0.643 and 0.343 mg/L, respectively. The DO values at the floating head, coma critical, coma, suffocation, suffocation, and death points of C. alburnus were 1.283, 1.063, 0.927, 0.863, 0.603, and 0.287 mg/L, respectively (Figure 2). In general, the DO values of floating head point, coma critical point, coma critical point, suffocation critical point, suffocation point and death point of C. alburnus were slightly higher than those of CM.

3.2.2. Effect of hypoxia stress on SOD enzyme activity of CM and C. alburnus
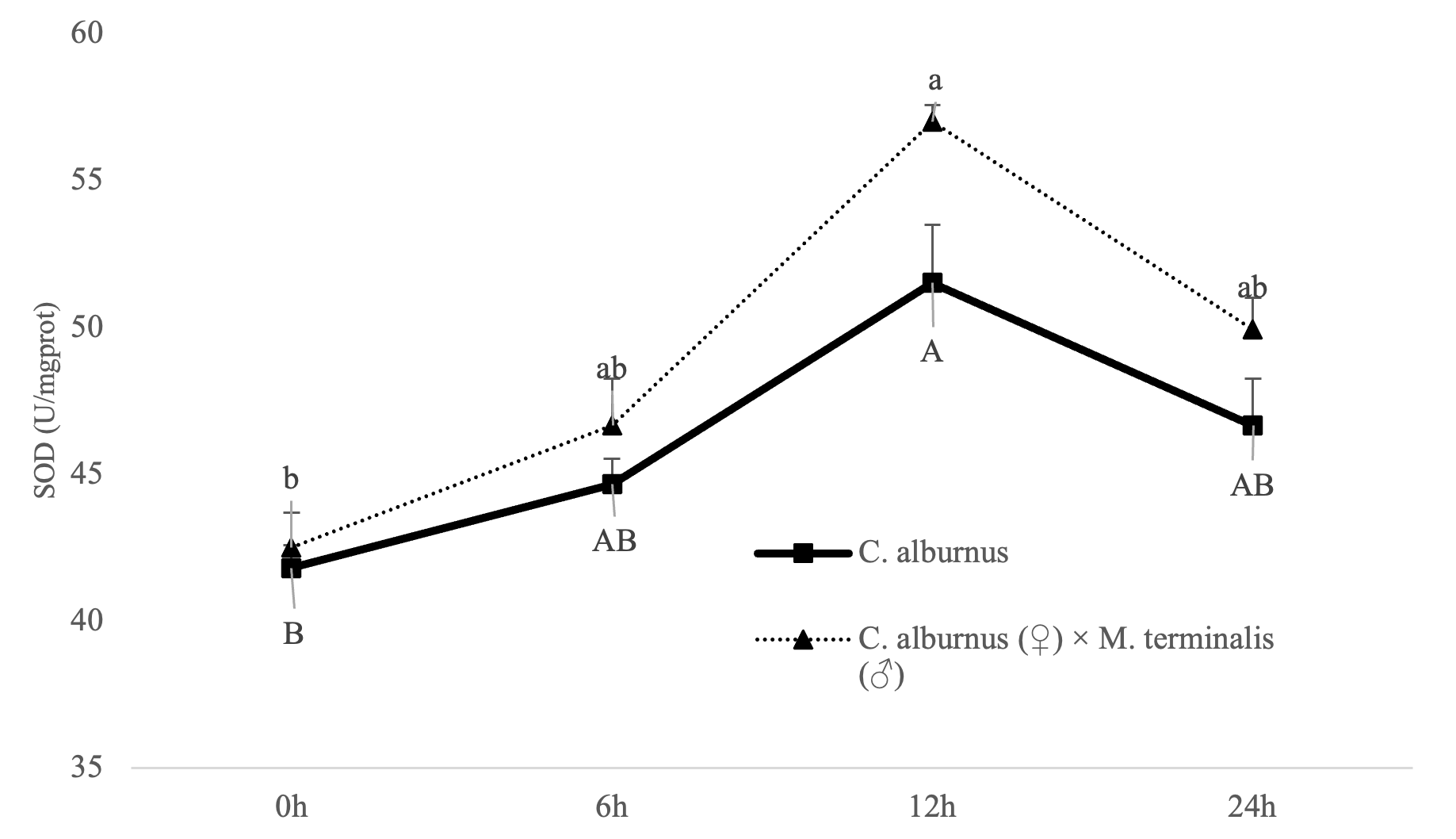
The change curve of SOD activity of CM and C. alburnus showed that SOD activity gradually increased at first and reached its maximum value at the 12th hour (SOD activity in C. alburnus was 52.53 U/mgprot, and SOD activity in CM was 55.83 U/mgprot). Afterwards, it decreased and remained higher than the value at 0 h after 24 h. Among them, SOD activity in CM or C. alburnus was significantly higher at 12 hours than at 0 hours (P<0.05) (Figure 3). In addition, when the SOD index of “All Female No.1” increased, the increase amplitude was greater, and when the activity decreased, the decrease amplitude was smaller. Overall, the SOD index of CM was more active than that of C. alburnus under the same hypoxia stress time.

3.2.3. Effect of hypoxia stress on GSH enzyme activity of CM and C. alburnus
The change curve of GSH activity of CM and C. alburnus showed that GSH activity gradually increased initially and reached its maximum value at the 12th hour. Afterwards, it decreased and remained higher than the value at 0 h after 24 h. Among them, GSH activity in CM or C. alburnus was significantly higher at 12 hours than at 0 hours (P<0.05) (Figure 4). In addition, when the GSH index of “All Female No.1” increased, the increase amplitude was more significant, and when the activity decreased, the decrease amplitude was smaller. Overall, the GSH index of CM was more active than that of C. alburnus under the same hypoxia stress time.

3.2.4. Effect of hypoxia stress on MDA activity of CM and C. alburnus
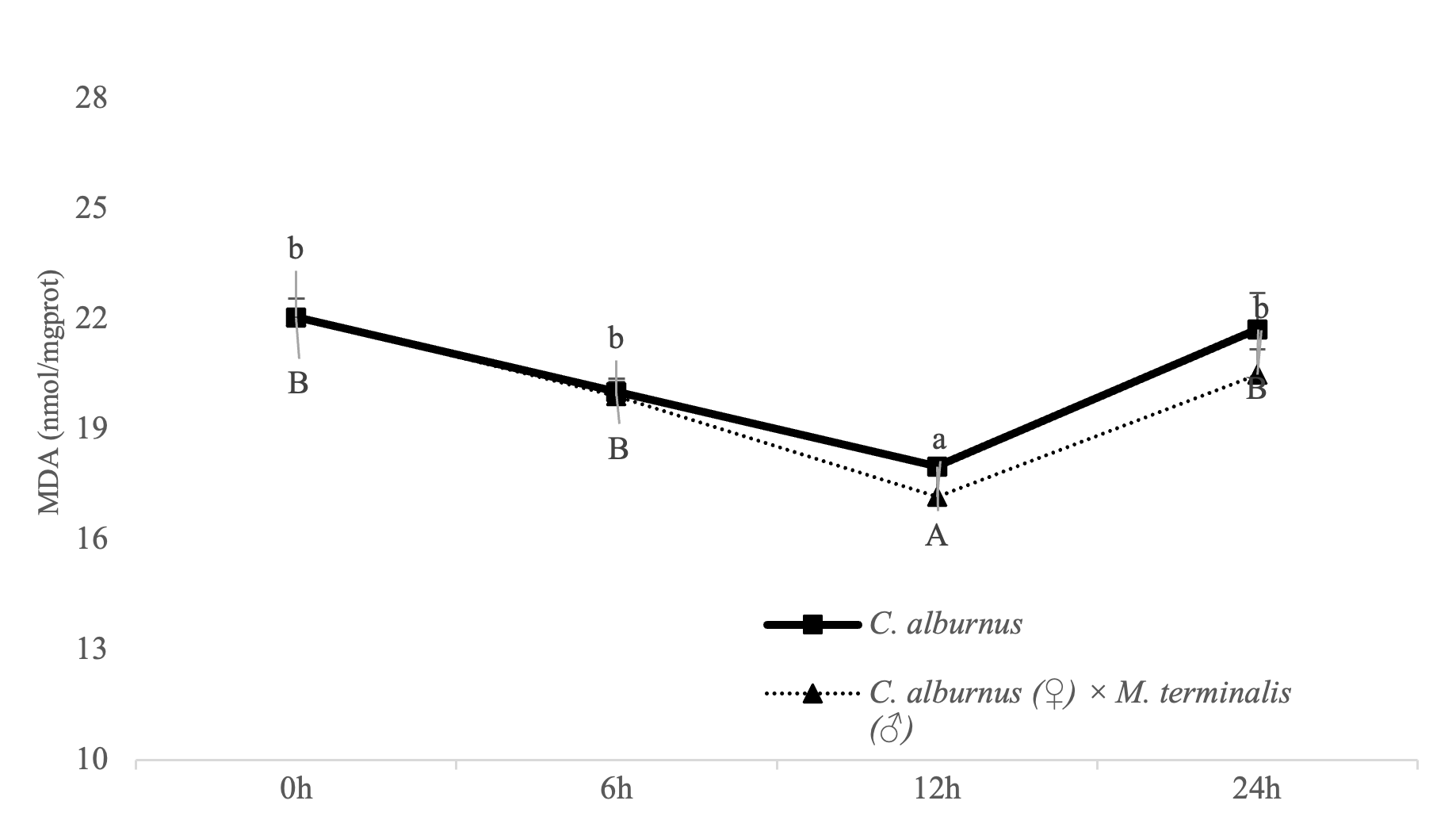
The change curve of CM and C. alburnus MDA activity showed that MDA activity gradually decreased at first and reached its minimum value at the 12th hour. Afterwards, it increased and remained lower than the value at 0 h after 24 h. Among them, MDA activity in CM or C. alburnus was significantly higher at 12 hours than at 0 hours (P<0.05) (Figure 5). In addition, when the MDA index of “All Female No.1” decreased, the decrease in amplitude was more significant, and when the activity increased, the increase in amplitude was smaller. Overall, the MDA index of CM was more active than that of C. alburnus under the same hypoxia stress time.

4. Discussion
4.1. Morphological characteristics between CM and Culter alburnus
Morphological proportional traits are essential indicators for fish classification.15 Guan et al.16 conducted a discriminant analysis of 29 proportional traits in seven populations, with a comprehensive discriminant rate of 86.3%. The discriminant effect was good, and morphological differences between different populations were observed. They found that the in the morphological characteristics of Megalobrama amblycephala, Magalobrame terminalis, Parabramis pekinensis, their hybrid offspring were very similar and difficult to distinguish with the naked eye. When analyzing the countable traits of the seven populations, it was found that the countable traits of the hybrid offspring were between the parents, and there were no significant differences, making it difficult to distinguish between the hybrid offspring. Zhao et al.17 showed that there were no significant differences in countable trait data of the hybrid F1 of M. amblycephala (♀) × P. pekinensis (♂) through a chi-square test. Li et al.18 found no significant differences in morphological traits in the genetic relationship analysis of M. amblycephala, M. terminalis, and Megalobrama hoffmanni. Chen et al.19 also found that the differences in countable traits among six Megalobrama populations in China were insignificant. Compared with previous studies, this study found that the main characteristics different from CM and Culter alburnus in appearance from Culter alburnus were the increase of body height and body thickness, the small blunt head, the bulge of the back of the head, and the large scales on the side of the body. Among the measurable traits, body length/height, body length/body thickness, tail handle length/tail handle height, and body length/gut length of CM were significantly smaller than those of Culter alburnus, while body length/tail handle length was significantly larger than that of Culter alburnus. The results showed that the morphological characteristics of CM were generally similar to those of Culter alburnus, but there were differences in body height, body thickness, tail handle, scales, etc. Especially the difference on body length/height and body length/body thickness, the critical morphological ratios showed a significant difference in the appearance of fish, whether they were long and thin or short and chubby. Studies have shown that distant hybridization of fish can effectively improve growth rates, transfer excellent traits, and cultivate hybrid offspring that integrate the superior genotypes of both parents, making it an easy breeding strategy for aquaculture breeders.20,21 Therefore, this study demonstrates the enormous potential of the hybridization of Culter and Megalobrama, more distant hybridization experiments could be designed to obtain rich breeding materials.
4.2. The hypoxia tolerance between CM and Culter alburnus
In this study, we determined the DO values at the floating head point, coma point, and suffocation point, finding that the values of CM were slightly higher than those of the ordinary Culter alburnus. However, based solely on these measurement data, it was difficult to determine that CM had a lower tolerance to low oxygen than ordinary Culter alburnus, because there was a small difference in indicators such as floating head point, coma point, and suffocation point between CM and Culter alburnus, and the weight of CM was slightly greater than that of ordinary Culter alburnus. Burleson et al.22 and Robb et al.23 studied stream fish and Micropterus salmoides and found that smaller fish species had stronger hypoxia tolerance than larger fish species. Therefore, it cannot be ruled out that slight differences in fish size might have led to differences in the results. Many new hybrid varieties have been obtained in China through distant hybridization between genera or above.24 The hybrid growth advantage of Magalobrama was more evident than that of its parents, and it significantly improved its tolerance to low oxygen levels and transportation compared to its parents.25 Zheng et al.26 found that an intermediate-type hybrid between the two species showed good growth advantages, with significantly higher total length, body length, and body weight indicators than the female parent Megalobrama amblycephala and the male parent Culter alburnus. CM in this study also had similar characteristics.
On the other hand, SOD, as an important antioxidant enzyme, could eliminate excess free radicals in organisms and was closely related to the immune ability of fish.27 It is generally believed that SOD plays a role in clearing reactive oxygen species earlier than other antioxidant enzymes. It first promotes the dismutation of O2- into H2O2 and O2, thereby detoxifying body.9 In this study, we observed that the liver SOD activity gradually increased under hypoxic stress, reaching a maximum at the 12th hour. Similarly, antioxidant substances such as GSH and MDA can reflect the health status of the body to a certain extent.28 Previous studies have shown that a decrease in GSH and an increase in MDA are not conducive to maintaining the dynamic balance of free radicals in the body, have a negative effect on normal physiological metabolism, and can easily lead to the development of diseases, thereby affecting survival status.28,29 The MDA content in this study first gradually decreased, reaching its lowest value at 12 h, and then increased.
Under low-oxygen stress, the activity of antioxidant enzymes in the body increases, inhibiting the production of oxidative free radicals. However, the activity of antioxidant enzymes in the body is inhibited under low oxygen stress, resulting in decreased antioxidant capacity. The body cannot eliminate the generated oxygen free radicals in a timely and effective manner, which accelerates lipid peroxidation and leads to an increase in the lipid product, MDA. This is similar to the conclusion of Lushchak et al.30 that fish may increase the activity of certain antioxidant enzymes in anaerobic or hypoxic environments. Cheng et al.12 reported that varieties with strong stress resistance showed a significant increase in antioxidant enzyme activity during adversity but a small decrease in activity during decline. Varieties with strong stress resistance showed increased activity, whereas those with weak stress resistance showed decreased activity. This study compared the changes in SOD, GSH, and MDA activities between CM and Culter alburnus in hypoxic stress experiments, finding that overall, CM had a higher ability to scavenge reactive oxygen species in hypoxic environments than ordinary Culter alburnus. Based on the measurement results of DO values at the floating head point, coma point, and suffocation point, we speculated that the tolerance of CM to hypoxic environments might be slightly higher than that of the ordinary Culter alburnus.
Based on the DO values, changes in SOD, GSH, MDA activities, and liver histological measurements of the floating head point, coma point, and suffocation point of CM, we speculated that the tolerance of CM to hypoxic environments might be slightly higher than that of the ordinary Culter alburnus. Prior literature on the genetic basis for hypoxia tolerance in hybrids had also reached similar conclusions.31,32 These results indicated that long-term hypoxic stress might cause oxidative damage to hybrid catfish and yellow catfish, but hypoxia tolerance of hybrid catfish was higher than that of yellow catfish. Further research was needed to determine whether it was due to the effects of methods such as coordinated reproduction between different families during the cultivation of CM. Simultaneously, in the actual production process, it is necessary to monitor the DO in the water in real-time to avoid the impact of long-term hypoxia on the body. It was necessary to further understand the response mechanism of CM under hypoxia stress in the future.
5. Conclusion
In this study, we selected CM and Culter alburnus to study their body shape characteristics and hypoxia stress. The results showed that CM was generally close to Culter alburnus in morphological characteristics, but there were differences in body height, body thickness, tail stalk, scales, etc.; At the same time, from the floating point to the death point, the DO value of Culter alburnus was generally higher than that of CM, and the SOD and GSH indexes of CM increased more when it increased, and decreased less when its activity decreased. On the whole, the SOD, GSH, and MDA indexes of CM were more active than that of Culter alburnus under the same hypoxia stress time. Therefore, it was speculated that CM had more robust hypoxia tolerance than that of Culter alburnus
Acknowledgements
This work was financially supported by the Cultivation and demonstration of new varieties of Culter Alburnus with high quality and stress resistance (2021C02069-3).
Authors’ Contribution
Conceptualization: Shun Cheng (Equal), Meifang Zhang (Equal), Meili Chi (Equal), Shili Liu (Equal), Miao Peng (Equal), Fei Li (Lead). Investigation: Shun Cheng (Equal), Meifang Zhang (Equal), Meili Chi (Equal), Jianbo Zheng (Equal). Formal Analysis: Shun Cheng (Equal), Wenping Jiang (Equal), Jianbo Zheng (Equal), Xiaoying Hang (Equal). Writing – original draft: Shun Cheng, Fei Li (Lead). Writing – review & editing: Fei Li (Lead). Supervision: Fei Li (Lead).
Competing Interest – COPE
The authors declare that there are no competing interests.
Ethical Conduct Approval – IACUC
This study was approved by the Ethics Committee of Laboratory Animal Center of Zhejiang Institute of Freshwater Fisheries. The animal protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of the Zhejiang Institute of Freshwater Fisheries. The animal study was reviewed and approved according to the guidelines of the Animal Experiment Committee, Zhejiang Institute of Freshwater Fishery (ZIFF20200304).
Conflict of interest
The authors declare that there are no competing interests.
Availability of data and material
The data and material are available by request.
Code availability
Software application.