1. Introduction
The Hippocampus abdominalis, commonly known as the pot-bellied seahorse, belongs to the family Syngnathidae within the order Gasterosteiformes. It is among the largest seahorse species and is primarily distributed in the marine waters of Australia and New Zealand in the South Pacific.1 In the aquaculture industry, seahorses are high-value marine species with significant trade volumes in markets such as traditional Chinese medicine and the aquarium trade. They hold an important position in these markets and are also one of the main species cultivated.2,3
Dissolved oxygen (DO) content in aquaculture water is a critical factor for fish survival, significantly influencing their energy consumption and metabolic rate.4 To enhance aquaculture efficiency and mitigate risks, extensive research has focused on the dynamic real-time monitoring of DO levels.5,6 Climate change-induced ocean warming and hypoxia have intensified oxygen stress for mariculture fish.7 Insufficient DO levels inhibit the growth of farmed fish, thereby reducing aquaculture productivity.8,9 Fish exhibit various adaptive strategies in response to differing DO concentrations in aquaculture water. When DO falls below 3 mg/L, fish are prone to disease and mortality.10,11 Temperature and DO are key environmental parameters in aquaculture, with DO closely correlate to water temperature.12–14 The oxygen consumption rate and suffocation threshold depend on multiple factors, including species, developmental stage, and external conditions such as water temperature, all of which influence fish survival and eco-physiological characteristics.15–18
In recent years, significant advances have been made in the domestication and breeding of H. abdominalis, with large-scale artificial breeding techniques becoming increasingly refined. However, current research primarily focuses on breeding technology,19,20 trait characterization,21,22 and genetic features.23 Studies indicate that temperature markedly influences the growth of H. abdominalis,1 yet data on how temperature affects oxygen dissolution and the suffocation threshold of this species remain limited. This study investigates the effects of different temperatures on DO content, oxygen consumption rate, and suffocation point in H. abdominalis, aiming to elucidate the basic parameters and principles of its respiratory metabolism. The findings are intended to inform physiological research, artificial breeding, and transportation practices for H. abdominalis.
DO concentration is a critical environmental factor in aquaculture, directly influencing the respiratory metabolism, growth, development, and survival of aquatic animals. Adequate DO levels support optimal physiological functions and provide a suitable habitat, while insufficient DO can cause hypoxic stress, growth inhibition, and even mortality.24 Numerous studies have demonstrated a significant negative correlation between water temperature and DO concentration: as temperature increases, DO concentration decreases. This phenomenon is primarily attributed to physical property whereby the solubility of gases in water declines with rising temperature. Consequently, higher water temperatures result in lower saturated DO concentrations.25–27
2. Materials and Methods
2.1. Experimental animals and acclimation
The experiment was conducted in November 2024 at the Changdao Enhancement Experimental Station of the Chinese Academy of Fishery Sciences. H. abdominalis specimens were obtained from Kunlunyangqi Marine Biotechnology (Shandong) Co., Ltd. A total of 150 healthy, uninjured individuals weighing (5.5 ± 1) g were selected and evenly divided into three groups of 50 individuals each.
During the experiment, the temperature of natural seawater was maintained at 14°C and adjusted using a heating plate. The temperature was increased or decreased by 2°C at each step, and the fish were acclimated for 1 hour before proceeding. Once the target temperature was reached, the fish were temporarily housed in four glass fiber reinforced plastic tanks, each with a diameter of 1 meter and a height of 1.5 meters. The tanks were set to water temperatures of 14°C, 19°C, and 24°C, respectively. The fish were kept in these tanks with gentle water flow for one week, with a temperature fluctuation range of ±1°C. The experimental water consisted of natural, sand-filtered seawater, which was aerated daily. The fish were fed frozen opossum shrimp to satiation twice daily, and uneaten food and debris were promptly removed. The fish were not fed 24 hours prior to the start of the experiment. During the experiment, the salinity and pH of the filtered seawater was monitored (YSI ProQuatro, USA), and the seawater salinity was regulated by manually adding freshwater or sea salt.The salinity of the experimental seawater was 28.25, and the pH was 8.54.
2.2. Test Methods
The static water respirometry method described by Dang Baocheng (2023) was employed, utilizing a 1.0 L cylindrical container (Songran SR-06) as the fish respiration chamber.28 Prior to the experiment, the chamber was placed in a water tank with a controlled temperature, ensuring that the container’s opening was slightly above the water surface. A water bath was used to maintain consistent temperatures between the chamber and the tank. A temperature range of 17-18°C was selected as optimal for the survival of H. abdominalis. This study aims to encompass the optimal temperature range for the H. abdominalis and investigate its DO tolerance under relatively low and high temperature conditions. The experimental temperature gradient was set at 14°C, 19°C, and 24°C, with temperature differences controlled and adjusted within 1℃. Each temperature test was conducted in triplicate, with a blank group serving as the control.
The fish were fasted for two days prior to the experiment to empty their digestive tracts and minimize interference from excreta. During the experiment, six fish were randomly selected from each group of fifty and placed into the respiration chamber. A probe from a portable water quality analyzer (YSI ProQuatro, USA) was positioned at the center of the bucket of water. The bucket opening and the cable were sealed with plastic wrap to isolate the air, and the initial DO concentration was measured. DO levels were recorded every six minutes under airtight conditions. Throughout the experiment, water temperature was monitored in real time, fish behavior was observed, and the time and DO concentration at which fish began to die, half of the fish died, and all fish died, were recorded. Upon the death of all fish, the experiment was terminated, and water on the fish surface was removed using filter paper. Body mass was then measured with an electronic analytical balance (Shanghai Jingke FA2004B). The body length was measured using a vernier caliper (SYNTEK JS20-GTG).
Formula for calculating oxygen consumption rate
R0=(ρn-ρn+1)×V/(m×t)
Where, R0 is the oxygen consumption rate [mg/(g·h)], V is the volume of the respiration chamber (L), ρn is the mass concentration of DO at the nth (n=1, 2, 3…) measurement (mg/L), m is the body mass of the fish (g), and t is the experimental time (h).
Determining the suffocation point: The concentration of DO when 50% of the experimental fish are observed lying on the bottom with their opercula no longer opening or closing is identified as the suffocation point for H. abdominalis at the specified experimental temperature.28
In this experiment, H. abdominalis demonstrated notable characteristics, including its relatively small size and upright swimming posture. It exhibited signs of distress earlier than anticipated, complicating the precise differentiation between behavioral stages. Consequently, we categorized the behavior of H. abdominalis in a hypoxic environment into two distinct stages: asphyxiation (Stage I) and death (Stage II).
Stage I encompasses the period from the onset of the experiment until the death of the first fish.
Stage II spans from the initial fish death to the mortality of all fish.
2.3. Data Processing
The experimental data were analyzed using SPSS version 20.0 software. Use a t-test to plot the data results as a composite line graph. Each data group was expressed as the mean ±standard deviation. A one-way analysis of variance (ANOVA) was conducted, followed by Duncan’s multiple comparison test to evaluate the differences between groups. A p-value of less than 0.05 was considered statistically significant.
3. Results
3.1. Dissolved oxygen content in water at different temperatures
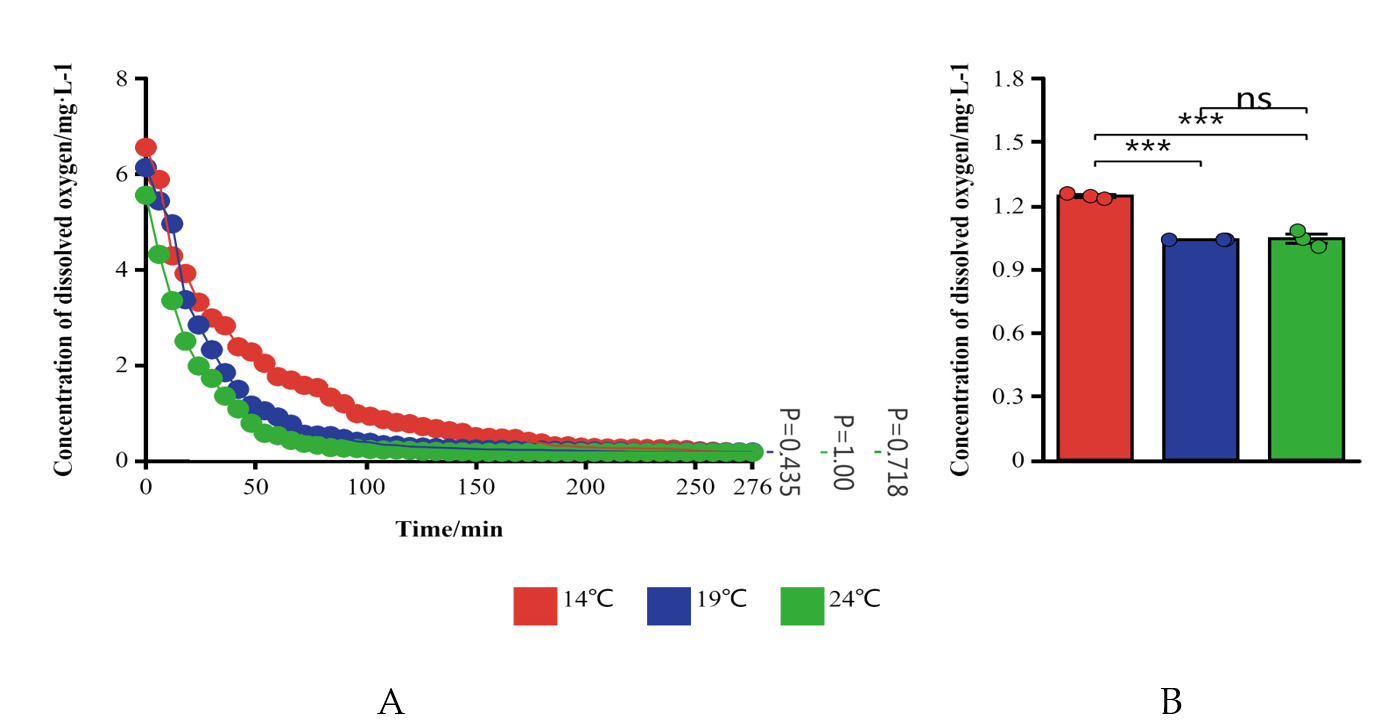
At the initial stage of the experiment, as the DO content in the water decreased, the opercular respiration of H. abdominalis became noticeably more intense, indicating a state of emergency stress (Fig. 1A). After a period, the subjects’ tails tilted upward to scratch their heads, and their heads swayed. As they adapted to the environmental conditions, their movements decreased, and they slowly sank, rolling onto their sides until they lay motionless at the bottom. Death was confirmed when both the tail and gills ceased movement.
The DO levels at the suffocation critical point, suffocation point, and death point of Haliotis abdominalis were measured across three temperature groups: 14°C, 19°C, and 24°C (Fig. 1B). The results were analyzed using a t-test and presented in a combined line and column chart.

The results demonstrated that the initial DO levels in the experimental groups at 14°C, 19°C, and 24°C were 6.55, 6.13, and 5.55 mg/L, respectively, indicating a decrease in initial DO content with increasing temperature. The time required for all H. abdominalis to perish in these groups was 276, 222, and 150 minutes, respectively. During the experiment, the DO levels in each group’s water initially declined and subsequently stabilized. At the conclusion of the experiment, the DO levels in the water were 0.16, 0.17, and 0.16 mg/L for each group, showing no significant change in final DO content with temperature increase. In the first 96 minutes, the differences in DO levels among the groups were statistically significant (P < 0.05), after which they stabilized with no significant differences. Overall, the average DO level in the 14°C group was significantly higher than in the 19°C and 24°C groups (P < 0.05), while no significant difference was observed between the 19°C and 24°C groups.
3.2. Oxygen consumption rates at various temperature combinations.
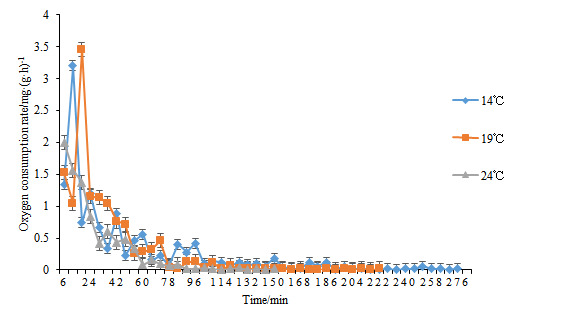
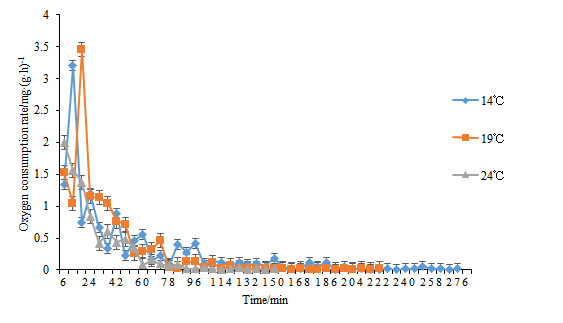
Table 1 demonstrates that oxygen consumption rates of H. abdominalis vary across different water temperatures. During the suffocation stage (Stage I), the rates were 0.29, 0.46, and 0.51 mg/(g·h) in the 14°C, 19°C, and 24°C groups, respectively. The rates in the 19°C and 24°C groups were significantly higher than in the 14°C group (P < 0.05). At the death stage (Stage II), the oxygen consumption rates decreased to 0.02, 0.01, and 0.01 mg/(g·h), respectively. Although the rate in the 14°C group was higher than those in the 19°C and 24°C groups, the differences were not statistically significant.
Figure 2 shows that over time, the oxygen consumption rates of H. abdominalis in the 14℃ and 19℃ experimental groups initially increase and then decrease, whereas in the 24℃ group, the oxygen consumption rate steadily declines. Under identical temperature conditions, as the DO content in the environment decreases, the oxygen consumption rate of H. abdominalis correspondingly declines. Notably, the 24℃ group reaches a low oxygen consumption state more rapidly than the 19℃ and 14℃ groups.
3.3. Suffocation points and behavioral performances under various temperature combinations
At the start of the experiment, the seahorses appeared relatively calm. As the DO concentration decreased, the fish gradually began to swim slowly. As the experiment progressed, the seahorses exhibited intermittent swimming and rapid head shaking. Ultimately, they became immobile, lying on their sides or backs, with their jaws open, and their dorsal fins and eyes ceased movement and rotation. They eventually died.
The suffocation points of H. abdominalis at each temperature were (0.18±0.03) mg/L, (0.20±0.05) mg/L, and (0.21±0.02) mg/L, respectively. The suffocation point increased with rising temperature, although there was no statistically significant difference among the temperature groups (Table 2).
At 14℃, DO decreased from 0.25 mg/L at the first death to 0.16 mg/L at the sixth death, a decline of 0.09 mg/L. At 19 °C, DO fell from 0.24 mg/L to 0.17 mg/L, a decrease of 0.07 mg/L. At 24℃, it decreased from 0.24 mg/L to 0.16 mg/L, a drop of 0.08 mg/L. Although the time to complete mortality differed across temperature groups, the overall reduction in DO concentration from the first to the last death was consistently close to 0.08 mg/L.
4. Discussion
4.1. Dissolved oxygen content and oxygen consumption rates in water under different temperature combinations
In this experiment, three distinct temperature groups exhibited differences in the initial mass concentration of DO. Dynamic monitoring revealed that, with increasing temperature, the rate of decline in DO concentration per unit time accelerated significantly across all groups. This observation suggests that higher water temperatures correspond to increased oxygen consumption by juvenile fish. Elevated temperatures enhance the metabolic activities of juvenile fish, thereby increasing their oxygen demand and accelerating DO depletion. These findings align with previous studies,28–30 which demonstrated that temperature influences the oxygen consumption characteristics of aquatic organisms.
At the end of the experiment, although the DO concentration in the 14℃ group was relatively higher than in the 19℃ and 24℃ groups, statistical analysis revealed no significant differences. This finding suggests that within the temperature range of 14-24℃, the oxygen tolerance of H. abdominalis tends to decrease as temperature increases, indicating a reduced ability to withstand hypoxia at higher temperatures. However, this temperature range may still fall within the species’ optimal adaptability. Additionally, factors such as sample size and measurement accuracy may have contributed to the lack of statistically significant differences in oxygen tolerance.
4.2. Effects of temperature on behavior, oxygen consumption rate, and suffocation point in Hippocampus abdominalis
Water temperature is a critical factor influencing biological growth, development, and energy metabolism. It significantly affects the oxygen consumption rate (OCR) and the suffocation point in fish. Previous studies have demonstrated that, within the temperature range of 13–28°C, the OCR of Hippocampus erectus increases progressively with rising temperatures, and the suffocation point similarly elevates.31 Consistent results were obtained in the present experiment for H. abdominalis within the temperature range of 14-24°C. In this study, the OCR in all groups exhibited a declining trend over time, while the OCR of H. abdominalis increased with higher temperatures. Over the course of the experiment, the OCR in each temperature group gradually decreased, and H. abdominalis displayed behaviors such as intermittent surfacing and head shaking. These observations suggest that the DO concentration in the water was insufficient, resulting in discomfort for the fish. Abnormal behaviors of this nature have also been documented in studies involving various other fish species.32 During this phase, H. abdominalis entered the asphyxiation period, during which the DO concentration in the water was insufficient to meet the juveniles’ basic oxygen requirements, inducing hypoxic stress responses. This observation aligns with the findings of Fu et al.,15 regarding the behavioral responses of fish under hypoxic conditions. In Stage I (the asphyxiation period), the OCR of all three experimental groups exhibited a fluctuating downward trend. The OCR in the 19°C and 24°C groups was significantly higher than in the 14°C group. However, no significant differences in OCR were observed among the temperature groups during Stage II (the death period). These results suggest that temperature during Stage I is the primary factor influencing OCR in juvenile fish. Furthermore, OCR decreased gradually across all temperature groups during Stage I, with significant differences observed, indicating notable variations in respiratory metabolism among juvenile fish subjected to different temperatures during the asphyxiation period.
In this experiment, the suffocation point of H. abdominalis increased with rising temperature. The suffocation point-the minimum DO concentration required for survival-indicates that higher temperatures elevate oxygen requirements and reduce oxygen tolerance in H. abdominalis. No significant difference in suffocation point was observed among the 14°C, 19°C, and 24°C groups, suggesting that small temperature changes (within 5 °C) in the 14–24 °C range do not appreciably affect the fish’s suffocation threshold. Within this range, H. abdominalis maintains dynamic equilibrium in oxygen utilization by increasing respiratory frequency at higher temperatures and reducing metabolic rate at lower temperatures, making fluctuations of ≤ 5°C insufficient to alter the suffocation threshold significantly. The suffocation point is defined as the DO concentration at which fish lose consciousness and approach death; in practice, it is measured as the DO concentration when 50% of the fish die. Our results also showed that the DO concentration at which H. abdominalis first exhibited signs of distress was substantially higher than its suffocation point across all temperature groups. These findings suggest that, despite hypoxia inhibiting respiratory metabolism, H. abdominalis retains the ability to mobilize energy under hypoxic conditions. This observation aligns with previous research in the field.33 Therefore, in aquaculture production, it is essential to monitor fish health and promptly detect DO levels in the water to prevent diseases caused by oxygen deficiency.
During the mortality period, the OCR of each temperature group increased with rising temperature. Higher temperatures shortened the time to mortality for all H. abdominalis individuals, with similar reductions in DO concentration across groups. For instance, at the three tested temperatures, the DO concentration decreased by approximately 0.08 mg/L from the first juvenile death to the death of all juveniles. In other words, following the initial juvenile death at different temperatures, a further decrease of 0.08 mg/L in DO concentration would result in the death of all juveniles. This phenomenon and its underlying mechanisms warrant further investigation.
The study found that at a water temperature of approximately 20°C, the OCR of H. abdominalis during the asphyxiation period was higher than that of Thamnaconus septentrionalis28 but lower than that of Pagrus major.34 However, the OCR during the death period and at the suffocation point was lower than that reported for other fish species. OCR and the suffocation point are influenced by multiple factors, including species, growth stage, body weight, and potential human errors in measurement. Consequently, the comparison results in this study may not fully capture the actual differences. Nonetheless, the findings suggest that H. abdominalis exhibits a higher OCR during the asphyxiation period, enters the discomfort phase earlier, and has greater oxygen demand in water.
5. Conclusions
The closed respirometer method was employed to investigate the variation patterns of OCR in H. abdominalis in relation to water temperature and DO concentration. This approach also facilitated analysis of its oxygen consumption capacity and oxygen tolerance. Results indicated that OCR increases with rising water temperature and decreases as DO concentration declines. Overall, H. abdominalis exhibits earlier signs of discomfort, requires higher DO levels, displays elevated OCR during asphyxiation, and enters a state of low DO and reduced oxygen consumption during death. The species demonstrates a narrow range of DO variation and limited DO tolerance. Insufficient DO can induce behavioral abnormalities in H. abdominalis, such as decreased locomotor activity, pronounced head shaking, and erratic swimming patterns. Therefore, particular attention should be given to the concentration of DO in water and its fluctuations during seed transportation and aquaculture operations.
Acknowledgments
We thank Runbo Zhang from Kunlun Yangqi Company for his help in sample collection. We also want to thank the reviewers for their constructive comments and useful suggestions, which improved the manuscript.
Author Contributions
Conceptualization: Chunnuan Zhao and Junjie Li.; Methodology: Chunnuan Zhao; Software: Chaoyi Xie; Formal analysis: Yang Chen and Liqun Ren; Resources: Bo Li, Yanxin Zheng and Nini Fan; Data curation: Yuping Wu and Shuai Xu; Writing - original draft preparation: Chunnuan Zhao; Writing - review and editing: Chunnuan Zhao and Tao Yu: Project administration: Chunnuan Zhao; Funding acquisition: Haiying Han and Tao Yu.
Funding
This work was supported by the Central Public-interest Scientific Institution Basal Research Fund,CAFS(NO. 2023TD69, Yantai Science and Technology Innovation Development Plan Project(NO. 2024YD084.
Ethical Conduct Approval – IACUC
The animal study was reviewed and approved by the animal care and use committee of Changdao Enhancement and Experiment Station, Chinese Academy of Fishery Science. All animal treatment protocols were performed in accordance with Chinese laws, regulations, and ethics.
Informed Consent Statement
Not applicable.
Data Availability Statement
Relevant information has been added in the article.
Conflicts of Interest
The authors declare no conflict of interest.