Introduction
In 2022, Mexico produced a total of 256,962 tons of shrimp, with 77% originating from aquaculture, 12% from estuarine, lagoon, and bay fisheries, and 10% from open-sea fishing.1 Shrimp farmers use sanitation measures, such as chemical applications to water and pond sediments. These measures are necessary to control and prevent the presence of disease-causing microorganisms.2 However, excessive use of chemicals could cause immunotoxicity and even reduce survival in farmed shrimp.
Shrimp defense system is composed of humoral and cellular effectors that together eliminate exogenous agents. Hemocytes are responsible for phagocytizing, encapsulating, and causing cytotoxicity. Three types of hemocytes are recognized: hyaline, semi-granular, and granular.3,4 Changes in total and/or differential number of circulating hemocytes in the hemolymph can be due to various factors, such as exposure to pollutants and physiological stress.5
Studies involving pesticide exposure have reported structural alterations in hemocytes and significant changes in various immunological parameters in shrimp. Additionally, these agrochemicals can reduce total protein content, elevate reactive oxygen species (ROS) production, induce lipid peroxidation, disrupt antioxidant enzyme activities (including catalase, glutathione peroxidase, and glutathione S-transferase), and cause DNA damage.6–8
Copper sulfate (CuSO4) is used in aquaculture to control macro and microalgal blooms and is also used in water to control the population of mussels (Mytilidae) in culture ponds.9 As observed in shrimp exposed to CuSO₄, normal hemolymph coagulation and hemocyanin concentration processes are disrupted, subsequently affecting respiratory function. Additionally, as reported, susceptibility to pathogenic microorganisms may increase under CuSO₄ exposure.2 A study on pesticide sales in northwestern Mexico revealed that insecticides accounted for 45.9%, herbicides for 30.5%, and fungicides for 20.1%, with most products containing metallic components.10 On the other hand, malathion residues of 0.0138 µg g-1 dry weight and copper sulfate residues (24 µg g-1 dry weight) were reported, both in shrimp from farms located in Northwestern Mexico.11,12 In this context, evaluating the effect of these chemical substances (malathion and copper sulfate) is important in the most popular shrimp rearing in the world.
As mentioned earlier, both chemicals can be found in shrimp ponds in NW Mexico; malathion presence in coastal water is due to agrochemical runoff, which is introduced to shrimp farms during pond water exchange, and copper sulfate is used for shrimp farming to reduce some organisms. The presence of both chemicals could affect shrimp physiological performance including immune response.13 Furthermore, the persistence of both chemicals in the pond water column may lead to their bioaccumulation in shrimp tissues. During effluent discharge, these compounds could be transported to coastal lagoons, raising concerns related to human safety and ecosystem health.
The present study is warranted because crustaceans are highly sensitive organisms whose health can be influenced by a wide range of chemical, physical, and biological changes in the aquatic environment. Consequently, immunological parameters serve as valuable indicators for assessing environmental health and quality. In Northwestern Mexico, the use of chemical agents in aquaculture is a common practice. Therefore, this study aims to evaluate and compare the effects of two substances—malathion and copper sulfate—on the cellular and humoral immune responses of shrimp when exposed individually to sublethal concentrations (below the LC50-96 h) under controlled laboratory conditions.
Materials and Methods
COLLECTION OF ORGANISMS
Juvenile L. vannamei were obtained from the “Los Sauces” shrimp farm (21° 35’ 54.6" N 105° 18’ 10.3" W) located in San Blas, Nayarit, México. 200 shrimp were transported to the laboratory in a 400-liter container. Thereafter, the organisms were acclimated to a salinity of 10 g L-1, pH of 7.8, a recorded oxygen level of 6.5 mg L-1, and an ambient temperature of 25 °C for three days to reduce transport stress. After this period, shrimp were randomly sampled and placed in 10-glass aquaria (60 x 40 x 33 cm) for each experiment performed (16 specimens/aquarium) and acclimated to these new conditions for three more days (habituation period).14
EXPERIMENTAL CONDITIONS
The experimental design was performed with two variations (concentration and time) sources in duplicate. The experiments were static and without water exchange; the animals were exposed for 96 h. A control group and four treatments were used for each of the chemicals tested. Malathion and copper sulfate concentrations were calculated from percentages (1, 10, 20, and 50%) of the median lethal concentration (LC50-96 h).9,15 The respective concentrations were 0.00078, 0.0078, 0.0156 and 0.039 mg L-1 and 0.35, 3.5, 7.02, 17.56 mg L-1 for malathion and copper sulfate, respectively. The test solutions were prepared as follows: 1000 mg L-1 stock solutions of malathion and copper sulfate (Baker, GR grade) in double-distilled water.
Before applying the chemicals, hemolymph samples were taken from shrimp (n = 4) from each aquarium, considering the control group (time 0). Once the different chemicals concentrations were applied, hemolymph samples were taken at 48 and 96 h.
HEMOLYMPH CLOTTING TIME MEASUREMENT
The hemolymph samples (10 μL) were taken with a 1-mL insulin syringe, which was inserted into the base of the first pereopod and placed in a 1.55-mm diameter hematocrit capillary microtube (BrandSD Germany). The time was recorded from the moment the needle was inserted and counted until the hemolymph stopped flowing through the capillary tube wall.16
HEMOCYANIN QUANTIFICATION AND GLUCOSE LEVEL CONCENTRATION
The hemocyanin concentration was determined from the hemolymph; 10 μL were taken in duplicate, poured into 1.5 mL cuvettes, and the sample was mixed with 990 μL of double-distilled water. Absorbance readings were taken on an Ultraviolet (UV)-10s VIS spectrophotometer (GENESYS-TM-USA) at a wavelength of 335 nanometers (nm), and hemocyanin concentration was reported in mmol L-1.17
For determining glucose level, 10-μL hemolymph was taken in duplicate and placed directly into the glucose meter (ACCU-CHEK® Active Roche DC México).18 Glucose dehydrogenase enzyme reacted in the test strip with the glucose present in the hemolymph. The subsequent chemical reaction changed the color of the test area and was quantified with reference value. The glucose value is expressed in mg dL-1 of hemolymph.
PHENOLOXIDASE (PO) ACTIVITY DETERMINATION AND TOTAL HEMOCYTE COUNT
The prophenoloxidase (proPO) and phenoloxidase (PO) activity were determined in triplicate in 30 μL of hemocyte lysate from each hemolymph sample on a 96-well microplate. Cacodylate buffer (30 μL), trypsin solution (100 μg/mL), and 170 μL of L-DOPA (L, 3,4-dihydroxyphenylalanine) (Sigma-Aldrich, USA) were used for the analyses. Absorbance readings were taken at 492 nm in an Awareness Technology microplate reader (Stat Fax® 2100, USA); PO activity is expressed in relative units (U PO mg protein-1).19,20
The total number of hemocytes was counted with an optical microscope (400X) and a Neubauer chamber, using four corner quadrants and an additional quadrant selected at random for each case. Total hemocyte counts were performed in 50 μL of hemolymph mixed with 50 μL of anticoagulant (27 mM trisodium citrate, 385 mM sodium chloride, 115 mM glucose, pH 6.6). Subsequently, 50 μL of the sample was mixed with an equal volume of formalin (37% formalin, 0.45 M NaCl), and the total hemocyte count was performed.
DIFFERENTIAL HEMOCYTE COUNT
For the differential hemocyte count, only the first 200 hemocytes were counted, and the results were expressed in percentages according to the three types of hemocytes that were observed, according to their size, shape, and granularity.
STATISTICAL ANALYSIS
The statistical values were analyzed with normality and homoscedasticity tests (Kolmogorov-Smirnov and Bartlett tests); subsequently, two-way analysis of variance (ANOVA) was applied when the statistical assumptions were fulfilled. Holm-Sidak comparison average test was also applied. The level of significance was P < 0.05 for all the tests.
Results
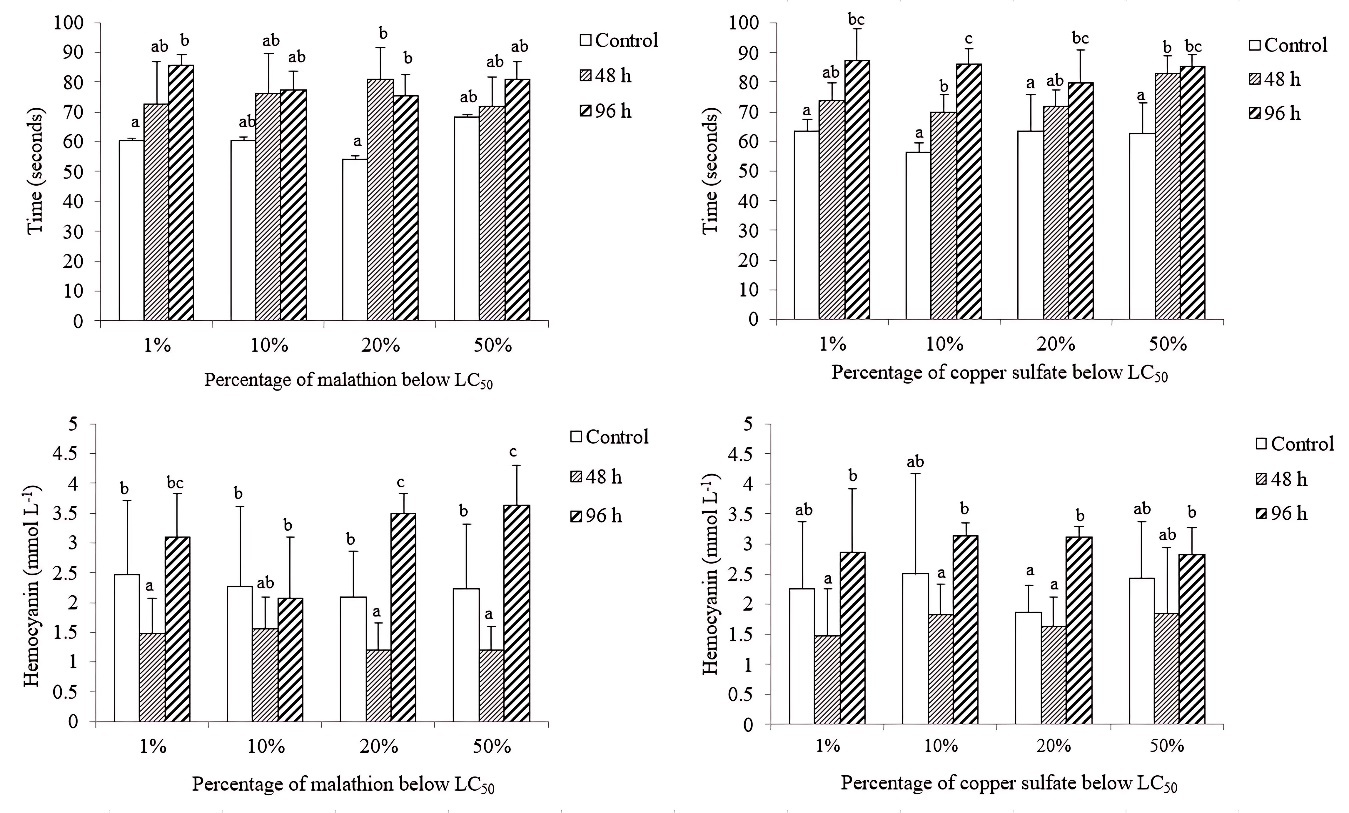
HEMOLYMPH CLOTTING TIME AND HEMOCYANIN CONCENTRATION
Concerning malathion exposure, 1% LC50 treatment caused 41% increase, while shrimp exposed to 20% LC50 treatment increased clotting time by 39%. In both treatments (96 h), the mean hemolymph clotting values were significantly different (P < 0.05) compared to the shrimp clotting time in the control group. For copper sulfate, the longest hemolymph clotting time was observed in shrimp in 1% LC50 treatment. Significant differences (P < 0.05) were found between the mean clotting times of shrimp exposed to the four treatments (LC50 1, 10, 20, and 50%), with respect to the average values of control shrimp. The greatest increase in coagulation time (54%) was present in shrimp exposed (96 h) to 10% of the LC50 of copper sulfate, reflecting a dose-dependent effect.
A reduction in hemocyanin content was also evident after a 48-h exposure, with significant differences (P < 0.05) among the average shrimp hemocyanin values from the malathion treatments (1, 20, and 50%) compared to the average shrimp hemocyanin values analyzed from the control group. While at 96 h exposure, the hemocyanin trend was to increase significantly (P < 0.05) in shrimp exposed to 20 and 50% of the LC50 of malathion, with respect to the average hemocyanin shrimp values from the respective control group.
In the four copper sulfate treatments, hemocyanin concentration decreased after the chemical application (48 h), but the percentage increased at 96 h exposure in shrimp exposed to the four treatments (LC50 1, 10, 20, and 50%) (Figure 1).
_of_hemolymph_clotting_time_(seconds)_and_hemocyanin_(mmol_l_-1_)_in_expo.jpeg)
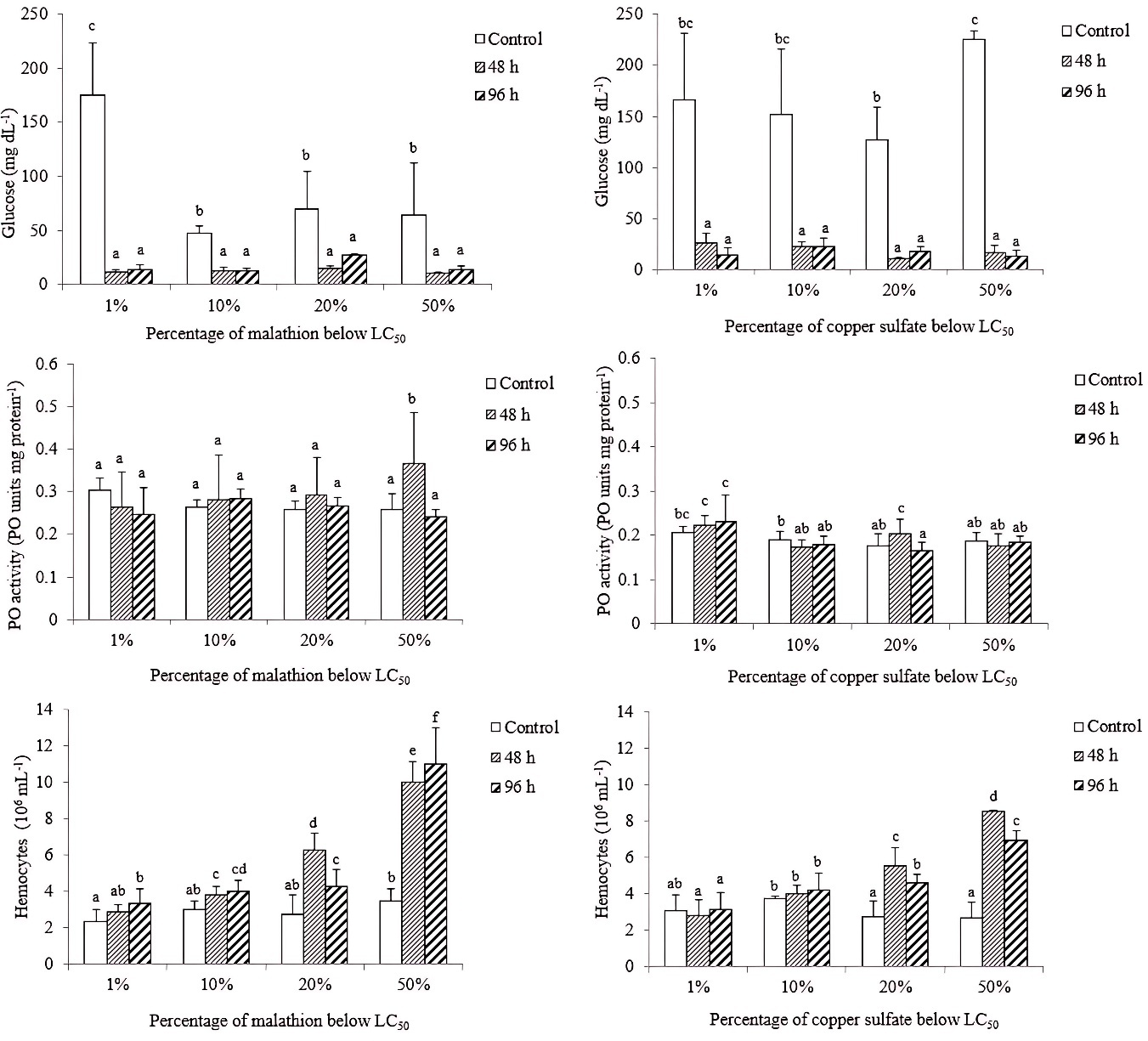
GLUCOSE LEVEL, PHENOLOXIDASE ACTIVITY (PO) AND, TOTAL NUMBER OF HEMOCYTES
Glucose levels were significantly lower (P < 0.05) in shrimp exposed to malathion and copper sulfate than those of the controls. However, no significant differences (P > 0.05) were registered among exposure times (48 and 96 h) for two experiments (malathion and copper sulfate).
For PO activity, only shrimp exposed to 48 h of 20 and 50% of the LC50 of malathion and copper sulfate were significantly different (P < 0.05) from those of the control. Concerning the total hemocyte count (THC), significantly (P < 0.05) higher values were determined in shrimp exposed to four malathion concentrations; while those shrimps exposed to copper sulfate, only specimens exposed to 20 and 50% of the LC50 were significantly higher than those of the control (Figure 2).
_of_glucose_(mg_dl_-1_)__po_activity_(po_units_mg_protein_-1_)_and_hemocy.jpeg)
DIFFERENTIAL HEMOCYTE NUMBER
The hyaline cell percentages decreased significantly (P < 0.05) according to time exposure with 78 and 69% lower than those specimens exposed to 20 and 50% (<LC50-96 h) of malathion, with respect to the control group. Regarding the hemocyte semi-granular (SG) average values, a significant increase (P < 0.05) was determined in shrimp (96 h exposure) at 20% LC50-96 h treatment with respect to the average SG value of the control group. Finally, the average number of granular hemocytes (G) tended to increase significantly (P < 0.05) as a time function in the organisms analyzed in all malathion treatments, compared to the shrimp G value of the control group (Table 1).
Regarding the copper sulfate bioassay, the different types of hemocytes showed a similar trend to that reported with malathion exposure. Hyaline hemocytes decreased by 74% in shrimp after exposure to 50% of the LC50-96 h of copper sulfate. Semi-granular hemocytes increased significantly (P < 0.05) as a function of time, except in the 20% LC50-96 h treatment. While mean granular hemocytes values increased at 96 h of copper sulfate exposure, the increases were approximately five times greater in shrimp in 20 and 50% of the LC50-96 h copper sulfate exposure, with respect to the values of the control group (Table 2).
Discussion
Hemolymph coagulation is an important factor in shrimp’s mechanism of response to avoid some diseases.21 In the present study, hemolymph coagulation of L. vannamei juveniles was altered when exposed to concentrations below LC50-96 h of malathion and copper sulfate. This observation is in concordance with other studies, where in coagulation time alteration of the crustacean hemolymph has been reported by pesticides, fungicides (Cu based), and the presence of microorganisms, such as viruses and bacteria.21,22 This effect could be caused by an alteration in the transglutaminase enzyme, which is responsible of the shrimp hemolymph coagulation time.23
Hemocyanin is a protein that transports molecular oxygen (O2) through the hemolymph to crustacean tissues, and changes in hemocyanin concentration have been reported and related to the effect of stress factors. In the present research, shrimp hemocyanin concentration caused a decrease at the first exposure time and an increase at the second exposure time below LC50-96 h of both substances tested. When the lobster Homarus americanus was exposed to the OP azamethiphos, an increase of hemocyanin concentration was observed. The authors pointed out that the increase could have been due to an increase in protein synthesis as a compensatory mechanism for the stress caused by chemical substances.24 An increase in hemocyanin concentration has also been reported in Cancer antennarius crab exposed to copper sulfate.25 This hemocyanin increase could be due to the stress caused by both chemical substances used.
Glucose level, as part of energy metabolism, plays an important role in shrimp homeostasis. A significant decrease in glucose level was determined in the present research after exposure to malathion and copper sulfate separately. An increase has been reported for L. vannamei and Macrobrachium nipponense in response to changes in water salinity.26,27 Furthermore, an increase in glucose level has been observed in Fenneropenaeus indicus exposed to malathion.28 This exposure altered glucose homeostasis, which has been reported as a possible toxicity mechanism associated with malathion exposure.29 The decrease in glucose levels observed in this study may be attributed to its utilization for ATP synthesis, resulting from elevated metabolic activity induced by the stress condition.30
The phenoloxidase (PO) is an important enzyme in the defense mechanism of crustaceans. In the present research, PO activity in shrimp exposed to malathion increased only in the 50% treatment of the LC50 at 48 h exposure time. In the shrimp Fenneropenaeus indicus the PO activity was altered when exposed to the OP malathion.28 Concerning copper sulfate, in the present study, PO activity increased in 20% and 50% treatments, with an LC50 at 48 h exposure; after that, PO activity tended to decrease in the indicated treatments at 96 h. The present results are consistent with Yuan et al.,31 who evaluated a diet containing copper sulfate plus copper amino acid in L. vannamei juvenile shrimp. The authors reported that PO activity in the hemolymph increased 3 h after the diet was included. For the Haliotis discus hannai an increase in PO was determined when exposed to different nickel concentrations.32 Since the proPO system is localized in hemocyte cells, when these cells are stimulated, the PO is released to hemolymph plasma,33 and it has a proportional relationship with the THC, which describes the same tendency.
A decrease in hemocytes can be caused by pigment synthesis, coagulation, wounds and even cuticle replacement functions, also due to protection against bacterial infections.34 Several factors, such as exposure to pollutants and physiological stress may cause the decrease in hemocytes.35 In the present study, L. vannamei juveniles were exposed to malathion, causing an increase in THC. The results in the present study agree with those reported by Chang et al.,36 when they used Macrobrachium rosenbergii species and were exposed to the organophosphate pesticide trichlorfon, after detecting an increase in hemocyte values in 12 and 24 h of exposure.
For the copper sulfate experiment, hemocyte tendency was to increase in number at 48 h and decrease at 96 h exposure to copper sulfate in L. vannamei. This result is in concordance with Roos-Muñoz et al.,37 who reported an increase in the total number of hemocytes in L. vannamei juveniles when exposed to mercury for 24 to 120 h. However, as per the report of Wei and Yang,13 the total number of hemocytes tended to decrease in Procambarus clarkii when exposed to copper sulfate at 48 h, indicating an interspecific variability.
Three types of hemocytes have been described, which can be classified according to their size, granule presence, and cytoplasm-nucleus ratio.38 In the present research, hyaline hemocytes of L. vannamei juvenile decreased with exposure to malathion and copper sulfate separately, and semi-granular and granular hemocytes increased after exposure for 96 h. Karamzadeh et al.38 experimented with L. vannamei fed with selenium and zinc nanoparticle diets and found that the most abundant hemocytes were in the order of hyaline, semi-granular, and granular, coinciding with that reported in the present study for juvenile L. vannamei exposed to malathion and copper sulfate separately. However, decreases in semi-granular and granular hemocytes are reported in L. vannamei exposed to zinc sulfate,39 which could indicate a metal-dependent effect. This decrease in hyaline cells could have been caused by an increase in phagocytic activity when these molecules (malathion and copper sulfate) are detected by the cell; however, future studies are required to provide a better explanation.
As a general conclusion, exposure to malathion and copper sulfate affected clotting time, glucose, THC, and PO activity in L. vannamei juveniles even below the median-lethal concentration. This result is of economic and ecological concern, since NW Mexico is an agricultural area of national importance. The presence of malathion and copper sulfate in shrimp aquaculture could have implications in aquaculture sustainability, food safety, and ecosystem health; and a mixture of these contaminants must be carried out in future experiments to know the specific effect of that mixture on L vannamei.
Acknowledgments
This research was funded by the National Council of Humanities, Sciences and Technologies (CONAHCYT) through National scholarship (1106141). The authors are grateful to Diana Fischer for the English edition.
Authors’ Contribution
Conceptualization: E.G. Hernández-Mendoza (Equal), Juan C. Bautista-Covarrubias (Equal). Methodology: F.J. Valdez-González (Equal), J.P. González-Hermoso (Equal). Formal analysis: E.G. Hernández-Mendoza (Equal), M. Aguilar-Juárez (Equal). Writing – original draft: E.G. Hernández-Mendoza (Equal), M.G. Frías-Espericueta (Equal), Juan C. Bautista-Covarrubias (Equal). Investigation: J.P. González-Hermoso (Equal), M. Aguilar-Juárez (Equal). Writing – review & editing: M.G. Frías-Espericueta (Equal), Juan C. Bautista-Covarrubias (Equal). Funding acquisition: Juan C. Bautista-Covarrubias (Lead). Supervision: Juan C. Bautista-Covarrubias (Lead).
Competing of Interest – COPE
Authors declare no competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical Conduct Approval – IACUC
The Bioethics Committee of the Autonomous University of Nayarit authorized this research.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.