Introduction
Abalone is one of the most valuable marine commodities globally, and its production has increased over the past decade owing to high demand. Overall abalone production, including that from both aquaculture and capture fisheries, has risen dramatically, from 14,599.00 tons in 2001 to 280,518.8 tons in 2023.1 The Pacific abalone (Haliotis discus hannai) is one of the most popular abalone species in East Asia, particularly in China and Korea. To remain sustainable, Pacific abalone production requires large quantities of high-quality sperm for use in in vitro fertilization.2
For organisms that practice external fertilization, reproductive cells are directly exposed to seawater, and environmental conditions significantly affect sperm quality and fertilization success. Changes in the chemical and physical properties of seawater can adversely affect sperm quality, with pH having a direct impact on gamete quality.3 As a result of ocean acidification, the pH of the Earth’s oceans is currently 0.1 units lower than it was since the Industrial Revolution, and it is anticipated to fall by 0.3–0.5 units by the end of this century.3 In coastal areas, seawater pH levels may fluctuate significantly, by up to 1 unit4; in some cases, it can decrease to as low as 6 or increase to as high as 10.5,6
Several studies demonstrate that pH influences sperm motility in marine organisms, including the Mediterranean mussel (Mytilus galloprovincialis), the ascidian (Ciona robusta),3 and the Manila clam (Ruditapes philippinarum),7 European eels (Anguilla anguilla),8 with specific effects varying among these species. Although the impact of seawater pH on sperm quality have been examined in various marine organisms, the impact of pH levels on Pacific abalone sperm remains unstudied. Comprehending these effects is essential for forecasting the pH tolerance threshold of Pacific abalone sperm and for managing potential future changes in Pacific abalone aquaculture. Sperm motility is the main indicator examined when assessing sperm quality, as it directly correlates with the success of fertilization. However, computer-assisted sperm analysis (CASA) enables the assessment of additional kinematic sperm characteristics, including average path velocity (VAP), straight-line velocity (VSL), curvilinear velocity (VCL), the amplitude of lateral head displacement (ALH), linearity (LIN), straightness (STR), and beat-cross frequency (BCF).8 The rate of oxygen consumption, which correlates with metabolic activity and the generation of energy for sperm motility, can be another useful indicator,9 and live/dead sperm viability can also serve as an indicator of fertilization success.2 An additional measure of sperm quality, DNA damage, can forecast potential reproductive dysfunction and embryonic developmental anomalies.10 Furthermore, oxidative stress indicators, such as hydrogen peroxide (H2O2) levels, can evaluate sperm quality, as elevated oxidative stress may result in sperm malfunction.11
Considering the importance of sperm quality in sustaining Pacific abalone production, the current study aimed to assess the influence of a wide range of seawater pH (6-10) on Pacific abalone sperm quality and fertilization success.
Materials and Methods
Sperm collection
Five adult male Pacific abalone (with a mean weight of 137.4 ± 2.24 g and mean length of 12.34 ± 0.11 cm) were acquired from a commercial abalone hatchery in Haenam-gun, South Korea. To induce spawning, we use the UV-irradiated seawater method. First, abalone with fully developed gonads were removed from water and air-dried while positioned in the shell-down and shell-up orientations for one hour each. Then, they were placed in UV-irradiated seawater with moderate aeration. The tanks were covered to simulate nighttime. Abalone was checked for spawning after one hour, then every 15-30 min for a further 2-3 h. After spawning, sperm suspensions were collected in Eppendorf tubes and promptly centrifuged at 4,000 rpm for 1 min. The supernatant was removed, and the sperm motility was observed using an optical microscope (CH30, Olympus, Tokyo, Japan) at 10× magnification. Using this method, the sperm motility remains above 80%.
Experimental design
This study aimed to investigate the impact of seawater pH on the quality of Pacific abalone sperm. We assessed five pH levels: 6.0, 7.0, 8.0, 9.0, and 10 for 15 minutes. Among these, pH values between 6 and 9 fall within the tolerance range of most aquatic species. pH 10 was selected to assess sensitivity beyond typical tolerance levels, as well as to account for high pH conditions in some coastal areas, particularly near disposal area6 and submarine groundwater discharge.12 In this analysis, pH 8 served as the control. The duration of pH treatment is selected according to the fertilization period between sperm and eggs of abalone, typically lasting for 15 minutes. The pH of artificial seawater (27 g L−1 of NaCl, 0.5 g L−1 of KCl, 1.2 g L−1 of CaCl2, 4.6 g L−1 of MgCl2, and 0.5 g L−1 of NaHCO3 in distilled water, DO 8.41 mg L−1, 34 PSU, 18℃) was modified using sodium hydroxide and hydrochloric acid. Sperm motility, progressive motility, kinematic parameters (VAP, VSL, VCL, LIN, STR, ALH, and BCF), DNA damage, H2O2 levels (H2O2), the oxygen consumption rate, live/dead sperm viability, acrosome integrity, and the fertilization rate were evaluated.
Motility, progressive motility, and sperm kinematic analyses
Sperm motility, progressive motility, and kinematic metrics, including VAP, VSL, VCL, LIN, STR, ALH, and BCF, were evaluated using a CASA system (Hamilton Thorne, Inc., USA). Sperm samples were mixed at a 1:100 (v/v) ratio in artificial seawater at different pH levels for 15 min, with five replicates for each pH value. The dilution values were chosen after adjustment to provide sufficient sperm for CASA while minimizing errors due to sperm collisions or too low sperm concentration. Then, diluted sperm samples were positioned on a 20 µm Leja slide (IMV Technologies, France) and assessed using the CASA software, the CEROS II instrument (Hamilton Thorne, Inc., Beverly, MA, USA) equipped with a Zeiss Axiolab 5 microscope at 10× magnification, including a CM-040GE camera with 0.4-megapixel resolution at 60 Hz for 30 number of frames (JAI, Tokyo, Japan). CASA settings included the following: exposure = 16 ms; gain = 300; minimum head brightness = 105; minimum head size = 3 μm2; maximum head size = 40 μm2; minimum tail brightness = 255, capillary correction = 1.3; maximum photometer = 80; minimum photometer = 25; minimum total cell count = 200; progressive STR = 45%; progressive VAP= 45 μm/s; slow VAP= 20 μm/s; slow VSL= 30 μm/s, static VAP= 4 μm/s, and static VSL=1 μm/s.
DNA damage assessment
DNA damage was evaluated using alkaline single-cell electrophoresis with CometAssay kits (Trevigen, USA), following the manufacturer’s guidelines. After a 15-minute pH treatment in artificial seawater, a sample was diluted in phosphate-buffered saline (PBS) to achieve a concentration of 1 × 106 sperm/mL. Then, the sample was blended with molten LMAgarose at a 1:10 (v/v) ratio, and 100 µL of the final mixture was promptly pipetted onto a CometSlideTM. The slides were then incubated for 10 min at 4°C in darkness, immersed in a lysis solution (2.5 M NaCl, 0.1 M EDTA, 10 mM Tris-HCl, 7.6 g L−1 NaOH pellet in distilled water) at 4°C in darkness for 30 min, and treated with an alkaline solution (200 mM NaOH, 1 mM EDTA) for an additional 30 min. Following these treatments, electrophoresis was conducted in an alkaline solution at 21 volts for 30 min, and the slide was subsequently soaked twice in dH2O, for 10 min each, and once in 70% ethanol, for 5 min. The specimens were stained using SYBR Gold (diluted 1:10,000 in DMSO) for 30 min and examined under a fluorescence microscope (DM 2500, Leica Microsystems, Germany) at 20× magnification for analysis. DNA damage was quantified as tail intensity from 100 individual sperm cells utilizing the Comet Assay IV Lite System (Instem, UK).
Hydrogen peroxide level measurement
Sperm samples were mixed at a 1:100 ratio in artificial seawater at various pH levels and incubated for 15 minutes. After the pH treatment, samples were centrifuged at 300 ×g for 5 min, and the supernatant was removed. Then, PBS was added to cleanse and gently resuspend the cells, and five hundred µL of the solution was homogenized at 4°C and maintained on ice until analysis. For analysis, samples were centrifuged at 4°C to remove insoluble materials, and a commercial kit, the Hydrogen Peroxide Colorimetric Assay Kit (Elabscience®, USA), was used to assess the H2O2 concentration in the supernatant. This kit can be used to measure H₂O₂ content in animal cell samples. The method is based on the reaction between ammonium molybdate and hydrogen peroxide, which produces a yellow color. The intensity of the resulting yellow color indicates the concentration of hydrogen peroxide in the sample cells. Additionally, the protein concentrations in the sperm samples were measured using the Bio-Rad Protein Assay Kit (Bio-Rad Laboratories, Inc., USA). The H2O2 and protein concentrations were quantified as absorbance at 405 and 595 nm, respectively, using a spectrophotometric microplate reader (SpectraMax 190, Molecular Devices, USA), with three replicates for each treatment.
Live/dead sperm viability and acrosome integrity measurement
The viability of sperm cells and the integrity of the acrosome were evaluated using the LIVE/DEAD® Sperm Viability Kit and LysoTracker Green DND-26, both acquired from Invitrogen Molecular Probes (USA). Sperm samples were diluted 1:100 in artificial seawater at a given treatment pH for 15 min. Following the pH treatment, the samples were centrifuged at 300 ×g for 5 min, and the supernatant was removed. A 10 mM HEPES, 150 mM NaCl, 10% bovine serum albumin buffer solution (pH 7.4) was utilized to wash and gently resuspend the cells. To assess cell viability, 5 µL of SYBR-14 and 5 µL of propidium iodide were mixed with 1,000 μL of the resuspended cell solution. To assess acrosome integrity, 5 µL of LysoTracker Green DND and 5 µL of propidium iodide were mixed into 1,000 µL of the resuspended sample. This yielded final concentrations of 100 nmol L−1 for SYBR-14 and LysoTracker Green DND and 12 µmol L−1 for propidium iodide. The final mixtures were gently vortexed and incubated in darkness for 15 min at 37°C, and the cells were promptly analyzed using a flow cytometer (Beckman Coulter, Inc., USA). The analyses were repeated to create three replications for each pH treatment.
Fertilization rate assessment
Five female abalone (with a mean weight of 188.3 ± 1.47 g and a mean length of 14.28 ± 0.12 cm) were transferred to inducing buckets containing UV-irradiated seawater with moderate aeration. Following spawning, egg quality was examined using a stereo microscope (Leica MZ10F, Leica Microsystems). Sperm and eggs were added at a 10,000:1 ratio to artificial seawater at variable pH levels, with three replicates for each pH level, and fertilization was allowed to occur for 15 min. Subsequently, three washes were conducted. The rate of fertilization (%) was assessed utilizing the formula:
Fertilization rate (%) = (Number of fertilized eggs / Number of counted eggs) × 100
Statistical analysis
Sperm quality indicators are expressed as means and standard deviations (SD). Statistical analyses were performed using SPSS version 25 software. Shapiro–Wilk tests were used to assess data normality, and Levene tests were employed to evaluate homogeneity of variance. One-way ANOVAs, followed by Duncan’s multiple-range tests, were conducted (p < 0.05) to analyze normally distributed data (Motility, progressive motility, VAP, VSL, VCL, damaged acrosome). For non-normally distributed data (STR, LIN, ALH, BCF. DNA damage, cell viability, and fertilization rate), Kruskal-Wallis tests, followed by Dunn’s tests, were conducted (p < 0.05).
Results
Motility, progressive motility, and sperm kinematics
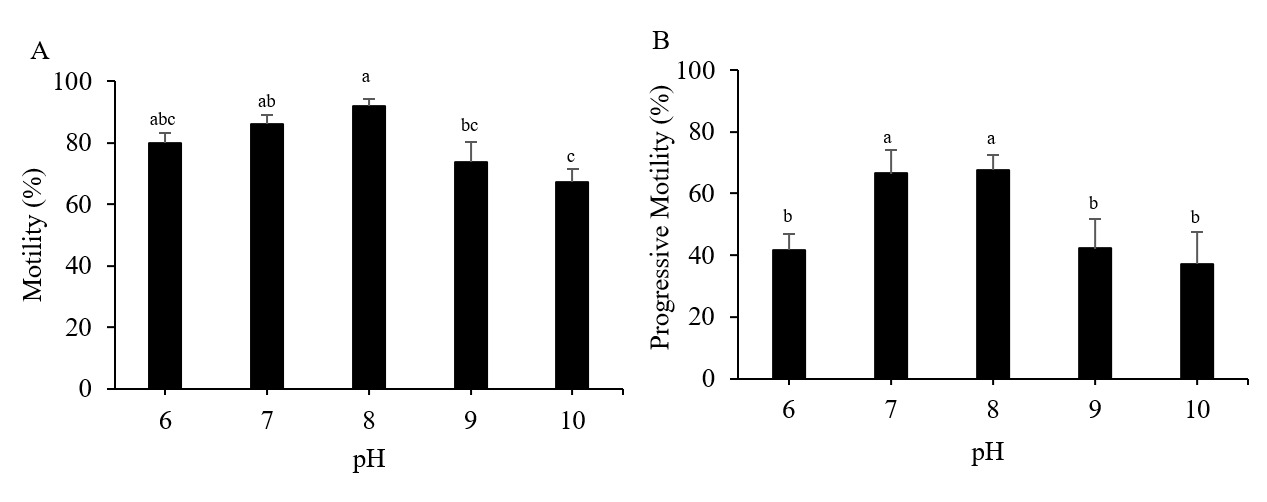
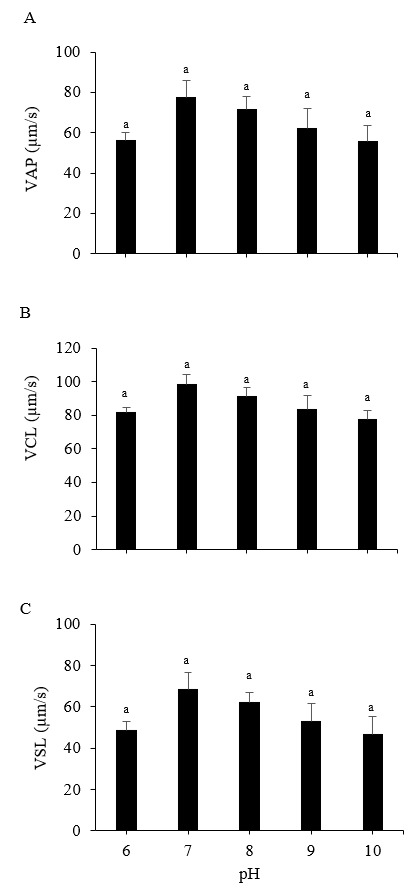
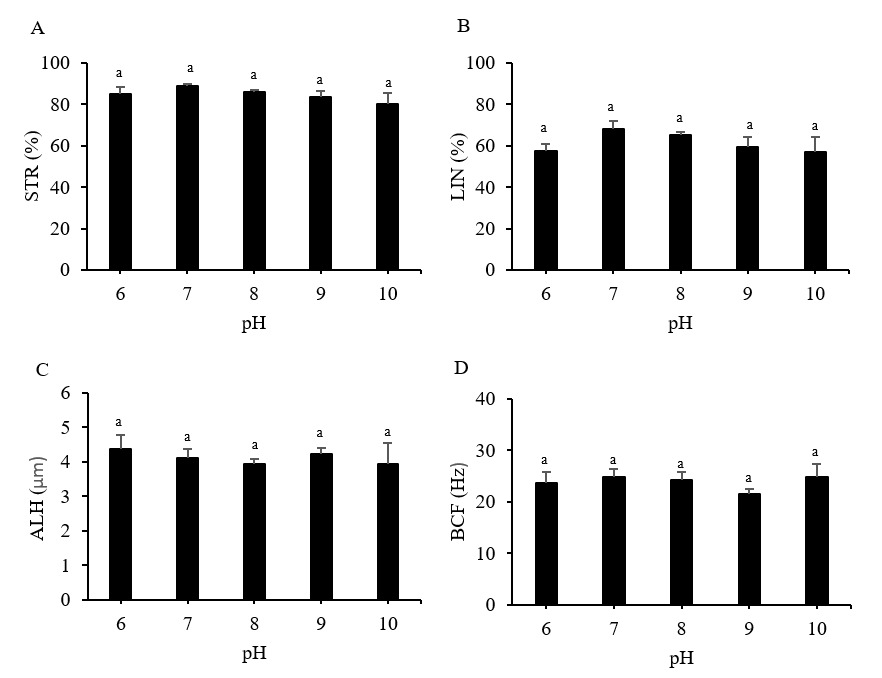
The Pacific abalone sperm remained motile in all treatment groups. However, motility was reduced significantly at pH 9 and 10. Whereas progressive motility was reduced significantly at pH 6, 9, and 10 compared to the control (pH 8) (p < 0.05). The greatest motility and progressive motility were achieved at pH 8, with values of 91.82 ± 2.46% and 67.54 ± 4.81%, respectively (Figure 1). The lowest values for both parameters were obtained at pH 10 (Figure 1). However, the values of the sperm velocity parameters, VSL, VCl, and VAP, under any pH treatment did not differ significantly from those of the control treatment (p > 0.05) (Figure 2). Furthermore, the pH treatments did not significantly impact the STR, LIN, ALH, or BCF values of Pacific abalone sperm (p > 0.05) (Figure 3).
_and_progressive_motility_(b)_of_pacific_abalone_(*halioti.png)
_sperm.png)
__linearity_(lin__b)__amplitude_of_lateral_.png)
DNA damage
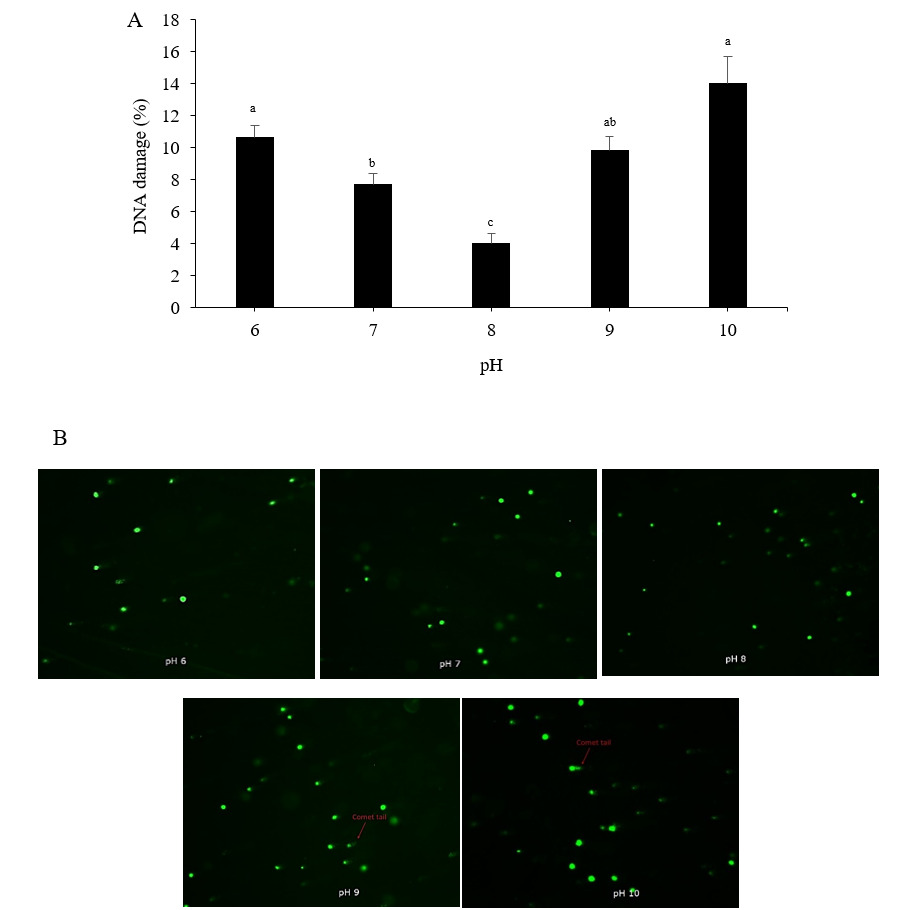
Tail fluorescence intensity can be used to estimate the amount of DNA damage. When compared to the control treatment (pH 8), exposure to low (pH 6), neutral (pH 7), and highly alkaline (pH 9 and pH 10) pH levels substantially increased DNA damage in Pacific abalone sperm (p < 0.05). The highest DNA damage level, 14.03 ± 1.68%, was observed when sperm were activated in pH 10 seawater, followed by the pH 6 (10.62 ± 0.76%) and pH 9 (9.82 ± 0.92%) treatments (Figure 4A). The lowest DNA damage value, 3.99 ± 0.66%, was recorded in the control treatment (pH 8). Figure 4B shows images illustrating the differences in fluorescence patterns among the pH treatments.
_sperm.png)
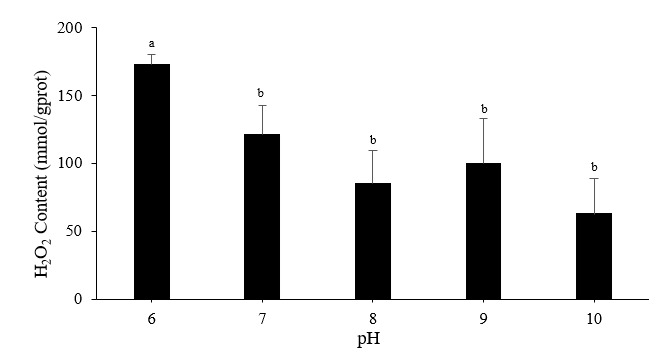
Hydrogen peroxide levels and oxygen consumption rates
The reaction between H2O2 and ammonium molybdate produced a yellow complex, which can be used to determine H2O2 levels. The acidic, pH 6 treatment significantly enhanced H2O2 production in Pacific abalone sperm, whereas the highly alkaline, pH 10 treatment did not alter the H2O2 level (Figure 5). The control pH induced the most excellent oxygen consumption rate (0.05894 ± 0.01912 nmol mL−1 min−1); there were no significant differences among the five pH treatments (p > 0.05).
_sperm.png)
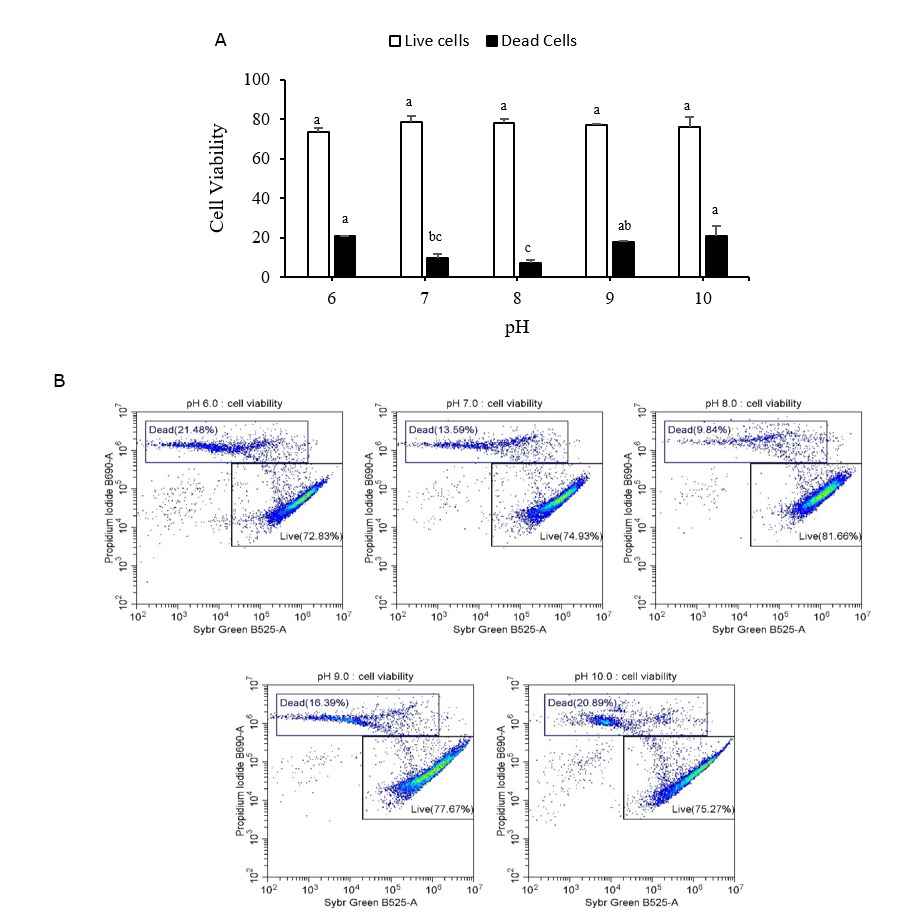
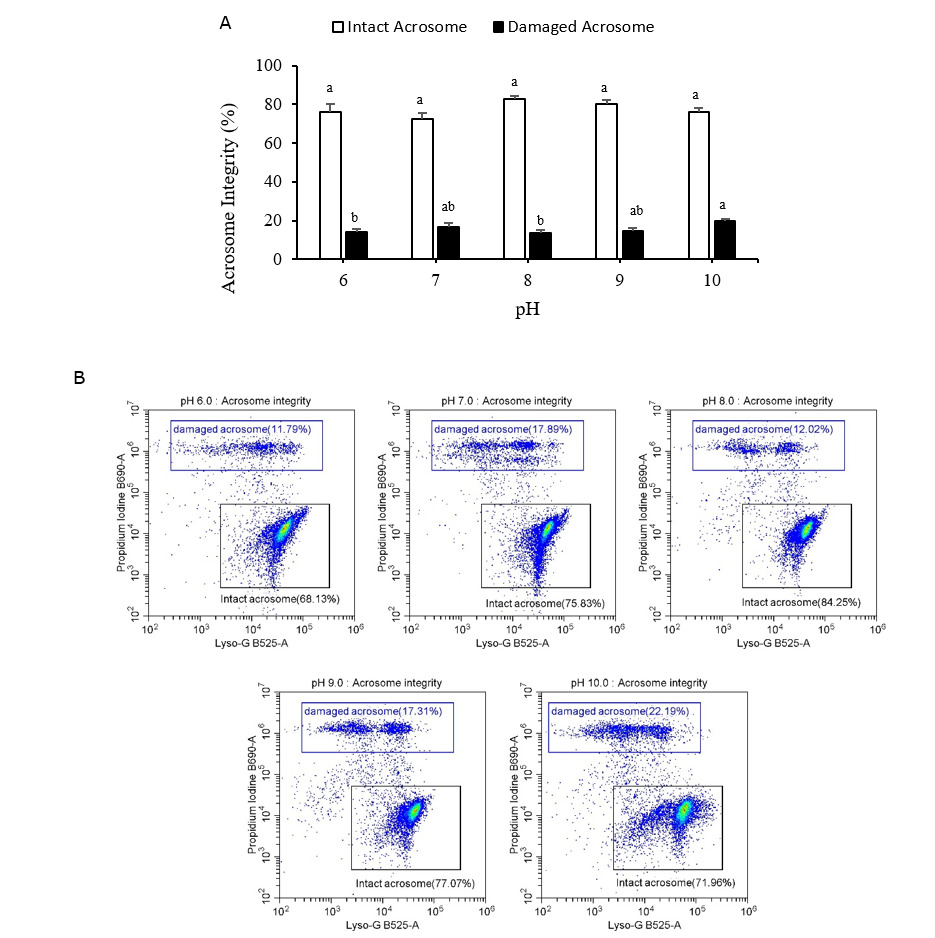
Live/dead sperm viability and acrosome integrity
Figures 6A and 7A display the live/dead sperm viability and acrosome integrity results, respectively. Both figures include representative flow cytometry output graphs (Figures 6B and 7B). Both indicators were measured using flow cytometry. The proportion of dead cells in the Pacific abalone sperm samples was significantly greater after 15 min of exposure to low (pH 6) or high (pH 9 and pH 10) pH levels than after exposure to the control-treatment (pH 8). The proportion of damaged acrosomes also increased significantly after exposure to pH 10 seawater. The pH 10 treatment resulted in the largest percentage of dead cells and damaged acrosomes (20.99 ± 5.01% and 19.97 ± 1.12%, respectively).
_sperm__me.png)
_sperm__m.png)
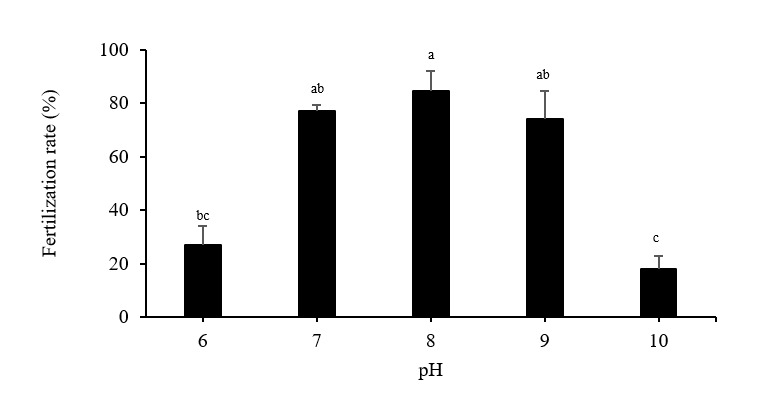
Fertilization rate
Fertilization rates fell significantly in the pH 10 and pH 6 treatments, which resulted in values of 17.83 ± 5.16% and 26.81 ± 7.18%, respectively (Figure 8). Fertilization rates at pH 7 (77.11 ± 2.19%) and pH 9 (74.08 ± 10.59%) were comparable to those seen in the pH 8 control treatment (84.56 ± 7.28%).
_fertilization_rates.png)
Discussion
Abalone sperm remains immotile in the gonads. Upon release into seawater or a diluent, their intracellular pH (pHi) increases,13 stimulating the activity of dynein ATPase, an enzyme involved in mitochondrial respiration, which in turn increases sperm motility.3 Seawater pH has been shown to impact sperm intracellular pH.3 In this study, Pacific abalone sperm motility was effectively initiated under all pH treatment conditions (pH 6–10), but progressive motility was reduced markedly at pH 6, 9, and 10. Decreases in sperm motility under low-pH conditions are assumed to be related to a drop in pHi, as has been demonstrated in the Mediterranean mussel, which exhibited decreases in both pHi and sperm motility after being exposed to acidified seawater for 30 or 60 min.3 This drop is further assumed to be caused by reductions in dynein ATPase activity, as this enzyme is known to function best in the 7.5–8 pH range.13 The availability of ATP, a crucial energy source, is also essential for sperm motility. As the power of flagellar movements increases, so do the energy requirements. Thus, a sufficient energy supply is crucial for sustaining motility over a suitable lifespan.9 Energy production in mitochondria may be reduced at low or high pH levels,14 meaning less ATP is available for use by dynein ATPase.
The optimal pH for initiating and achieving maximal sperm motility varies among species.7 Optimal pH values for sperm motility in the eastern oyster (Crassostrea virginica) range from 7.5 to 10,.15 Using artificial seawater, the optimal pH values for sperm motility were established as >7 for the Japanese scallop, >8 for the Pacific oyster, and >9 for the Manila clam.7 The composition of the activation solution may also influence the impact of pH on sperm motility. In a study using Coruh trout (Salmo coruhensis) and rainbow trout (Oncorhynchus mykiss), the optimal pH for sperm motility varied depending on the activation solution used. Notably, sperm motility for both species significantly declined when subjected to a pH 6 treatment in all activation solutions.16 Based on the results of the current study and the earlier study, we can see that the effect of the pH on the aquatic organism sperm is influenced by several things, namely species specificity, activation solution, and duration.
In the current study, neither velocity nor other CASA parameters (STR, LIN, ALH, and BCF) differed significantly among the pH treatments. Upon activation, spermatozoa show a linear or slightly curved trajectory. At the end of the activation stage, exposure to contaminants or an inappropriate extender may result in a progressively curved trajectory that ultimately leads to the formation of tight circles. Under these conditions, LIN [the ratio of net distance traveled to total path distance (VSL/VCL)] or STR [the ratio of net distance traveled to smoothed path distance (VSL/VAP)] can function as important parameters for assessing trajectory curvature.17 This suggests that while extreme pH treatments may not directly impair the individual motile sperm’s ability to enter and fuse with the oocyte, they may reduce the quantity of sperm reaching the egg by lowering the proportion of motile sperm.18
Any alteration of DNA that changes its coding properties or normal function in transcription or replication is referred to as DNA damage.19 Consequently, assessing DNA damage in sperm is crucial, as it impacts sperm quality, increases genetic and epigenetic abnormalities, and diminishes the sperm’s ability to produce normal embryos.10 The current study found that DNA damage increased with exposure to pH levels above or below that of the control, with maximum damage levels obtained at the extreme pH values of 10 and 6. Under acidic conditions and elevated temperatures, lambari (Astyanax altiparanae) sperm also exhibited increased DNA damage.20 The DNA helix is destabilized and DNA’s melting point is altered by extreme pH levels, regardless of whether they are low or high.21 Compared to somatic cells, spermatozoa possess restricted DNA repair mechanisms owing to the substitution of histones with protamines during spermatogenesis. Sperm cells are also susceptible to lipid peroxidation due to the high polyunsaturated fatty acid (PUFA) content of their membranes. Together, these factors render spermatozoa particularly vulnerable to oxidative damage.22 Despite a demonstrated correlation between environmental stress and DNA damage, rainbow trout (Oncorhynchus mykiss) sperm are still capable of accomplishing fertilization due to the ability of oocytes to repair DNA. Nevertheless, if DNA damage is excessive, fertilization failure may occur if the oocyte’s repair capacity is insufficient.23
Hydrogen peroxide is one of the most reactive oxygen species generated in sperm,24 and excessive H2O2 levels can adversely affect sperm cells, inducing oxidative stress.22,25 Nevertheless, sperm–egg fusion, chromatin stability, hyperactivation, acrosome reaction, and mitochondrial integrity all require a sufficient H2O2 level. The current study showed that H2O2 levels were substantially higher at pH 6 than at pH 8, indicating that Pacific abalone sperm experience increased oxidative stress at pH 6. Notably, H2O2 concentrations in sperm subjected to the extreme alkaline pH of 10 were the lowest among the treatments and did not differ substantially from those in sperm subjected to the control conditions. At high pH levels, H2O2 synthesis in cells can be impeded, primarily due to the impact of pH on the activity of the enzymes involved in the process. Additionally, at elevated pH levels, H2O2 can undergo dissociation.26 Overall, research into the impacts of pH on H2O2 levels in mollusk or invertebrate sperm using a wide pH range, such as 6–10, has been limited. However, a study looking at decreasing pH levels in the limited range compatible with current ocean acidification scenarios found that exposure to pH 7.8 for 30 or 60 min did not impact H2O2 levels in Mediterranean mussel and ascidian sperm.3 This result was attributed to an increase in antioxidant production that offset any increase in reactive oxygen species production.3 In another study, a pH of 7.7 also had no significant influence on intracellular H2O2-derived reactive oxygen species levels in Mediterranean mussel sperm.27
The proportion of dead Pacific abalone sperm cells and sperm with damaged acrosomes also increased at low or high pH levels, but for damaged acrosomes, this effect was only significant at pH 10. Under environmental conditions that exceed tolerance limits, reactive oxygen species production may increase, resulting in increased lipid peroxidation in the plasma membrane and the formation of lipid peroxides.25 The buildup of these lipid peroxides in the plasma membrane can cause cell death.27 To successfully enter the egg’s outer envelope and fuse with its plasma membrane, a sperm must execute the acrosome reaction, and intracellular pH affects the acrosome reaction. Intracellular pH changes away from the ideal level can reduce fertilization success by modulating the acrosome response.28 Furthermore, external pH can affect the internal pH of the egg, which is another important factor affecting a sperm’s ability to penetrate the egg cell.28 Consequently, fertilization rates were much lower at extremely low and high tested pH levels due to a decreased number of live cells and intact acrosomes. In blood clams (Tegillarca granosa), fertilization rates at pH 7.4 were greatly reduced compared to those of the control group,28 and the fertilization rates in the thick-shelled mussel (Mytilus coruscus) were considerably lower at pH 7.3 than at pH 8.1 (the control level).29 Another study subjected Far Eastern catfish sperm and eggs to pH values ranging from 2 to 13 during fertilization. Eggs could be fertilized in a pH range of 3–12, but fertilization rates decreased significantly at pH levels of 3, 10, 11, and 12, with no significant differences recorded between pH 4 and pH 9.30
The findings elucidate the tolerance capabilities of Pacific abalone sperm to pH using innovative technological methodologies, specifically CASA, comet assay, and flow cytometry. The data can be used to optimize hatchery conditions during spawning season and monitor seawater pH within the optimal range.
Conclusions
To summarize, Pacific abalone sperm motility can be initiated in seawater at pH levels in the 6–10 range. However, pH levels of 6, 9, and 10 can have a detrimental influence on abalone sperm quality, reducing motility and progressive motility while increasing the proportion of dead cells, and cells exhibiting DNA damage and damaged acrosomes. Fertilization might also be hindered at pH levels of 6 and 10. Future research into the combined effects of pH and other parameters is necessary, as numerous factors influence coastal seawater conditions.
Acknowledgments
This research was supported by the Korea Institute of Marine Science and Technology Promotion (KIMST), funded by the Ministry of Oceans and Fisheries (RS-2022-KS221671).
Authors’ Contribution
Conceptualization: Dian Y. Pratiwi (Lead). Methodology: Dian Y. Pratiwi (Lead). Formal Analysis: Dian Y. Pratiwi (Equal), Tae M. Kim (Equal), Young D. Oh (Equal), Mi-Jin Choi (Equal). Investigation: Dian Y. Pratiwi (Equal), Tae M. Kim (Equal), Young D. Oh (Equal), Mi-Jin Choi (Equal). Writing – original draft: Dian Y. Pratiwi (Lead). Writing – review & editing: Young D. Oh (Equal), Mi-Jin Choi (Equal). Funding acquisition: Kyu L. Han (Lead). Supervision: Kyu L. Han (Lead).
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
No ethical conduct approval-IACUC was required for this research.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.