Introduction
Water temperature is a significant environmental stressor affecting the physiological and biochemical state of aquatic animals.1 As typical poikilotherms, fish exhibit body temperature fluctuations closely linked to environmental water temperatures.2 When water temperature undergoes drastic fluctuations beyond the fish’s tolerance range, their health is significantly affected.3 Acute thermal stress has been shown to adversely affect fish. For instance, elevated temperatures significantly inhibit the growth of Oncorhynchus mykiss and impair intestinal structure and barrier function, leading to pathological damage.4 Acute high-temperature stress for 24 hours caused severe hemorrhaging in the livers of Micropterus salmoides.5 Furthermore, the abrupt rise in water temperature altered serum Glu, TP and ALB levels,6 disrupting energy metabolism and impairing immune function.
Blood serves as a vital medium for the transport of materials, metabolic regulation, and immune defense in fish. Changes in serum biochemical indicators provide a direct reflection of the physiological status and adaptive mechanisms of fish under high-temperature stress, making them crucial metrics for evaluating fish physiology, health, and environmental adaptability.7 The serum levels of AST, ALT, and Glu in Takifugu obscurus significantly increased,8 while ALP and TP levels significantly decreased under high-temperature stress. Additionally, cell viability was significantly suppressed and immune function declined; In studies of Labeo rohita,9 serum ALT and AST activity increased, while TP content decreased. During heat stress in Phoxinus lagowskii,10 the level of WBC, RBC, HGB, and HCT exhibited a trend of initially increasing followed by decreasing; Pangasianodon hypophthalmus exhibited a significant reduction in RBC and HGB counts after 3 and 7 days of exposure at a water temperature of 36°C, while WBC counts showed an opposite trend.11 Studies on Oreochromis niloticus12 revealed that RBC and HGB counts decreased significantly with rising temperatures, whereas WBC counts showed the opposite trend. At 7 and 15 days, Glu levels increased significantly with elevated temperature. This rise in Glu levels indicates enhanced glucose metabolism, leading to increased glycolytic pathway activity and a surge in reactive oxygen species (ROS). This process adversely affects immune function and antioxidant status. The activities of superoxide dismutase (SOD), catalase (CAT), and lysozyme (LYZ) in juvenile hybrid yellow catfish (Pelteobagrus fulvidraco × Pelteobagrus vachelli)13 initially increased but decreased after 48 hours at 35°C, The production of ROS in Argopecten irradians14 significantly increased, accompanied by elevated SOD and CAT activities, and upregulation of heat shock protein 70 (HSP70) after 6 hours of heat stress. Malondialdehyde (MDA) levels peaked at 12 hours, while SOD and CAT activities declined. Therefore, elucidating the physiological mechanisms underlying fish responses to environmental stress has significant practical implications for advancing the scientific and healthy development of fish farming.
Blunt snout bream, Megalobrama amblycephala, is one of the key freshwater aquaculture species in China15 and is characterized by its excellent meat quality, rapid growth rate, and high survival rate.16 According to the China Fisheries Statistical Yearbook, in 2022, China’s total farmed production of crucian carp and grass carp reached 767,000 metric tons (with blunt snout bream accounting for over 90%). The annual output value of the entire industry chain amounted to approximately 45 billion yuan, ranking eighth among freshwater farmed fish species in China. Currently, there is limited research on the physiological responses of M. amblycephala to high-temperature stress, primarily focusing on changes in physiological and immune indicators during chronic heat stress or at single time points. However, systematic studies on hematological and plasma biochemical responses at different time points during short-term high-temperature stress remain relatively scarce. In particular, the physiological adaptation mechanisms and recovery potential of M. amblycephala during stress progression under 34°C high-temperature conditions remain unclear. Therefore, this study selected 25°C as the control group and 34°C as the high-temperature treatment group. We investigated the effects of acute high-temperature stress at 6 different time points (0, 3, 6, 12, 24, and 48 hours)17 on plasma biochemical and hematological parameters in M. amblycephala. The aim is to reveal the physiological temporal response patterns of M. amblycephala under short-term high-temperature stress, elucidate the dynamic mechanisms of its metabolic regulation and immune adaptation, and fill the temporal gap in systematic physiological data for this species during short-term heat stress. To systematically examine the physiological adaptation mechanisms of M. amblycephala to heat stress, this study proposes the following specific hypotheses based on the common effects of high temperatures on fish metabolism, liver function, and hematology: Heat stress may cause liver dysfunction or liver damage in M. amblycephala; heat stress may affect the physiological metabolism, immune capacity, and blood oxygen-carrying capacity of M. amblycephala. By validating these hypotheses, this study not only provides time-dependent physiological data to understand the heat-adaptation mechanisms of M. amblycephala but also provides a basis for early temperature warning and management strategies in practical aquaculture. It holds significant practical implications for advancing the sustainable farming of M. amblycephala.
Materials and Methods
Experimental fish
The juvenile M. amblycephala used in the experiment were sourced from the Freshwater Fisheries Research Center of the Chinese Academy of Fishery Sciences. The experimental fish were acclimated for 14 days in cylindrical water tanks (100 L/tank). During acclimation, they were fed twice daily with commercial feed (Wuxi Tongwei Feedstuffs, Wuxi, China) containing 40% protein and 5% fat until satiated (7:00 AM, 6:00 PM, not exceeding 3% of body weight), water exchange rate was 33% every two days, to adapt to environmental conditions prior to the acute high-temperature exposure experiment. Additionally, feeding was suspended for 24 hours prior to the experiment.
After acclimatization, we selected 120 healthy M. amblycephala of uniform size, with an average weight of (17.72 ± 0.05 g). Summer water temperatures typically exceed 30°C. Research has indicated that 34°C can induce heat stress.18 Therefore, the experiment was divided into a control group maintained at (25.25 ± 0.34°C) and a treatment group at (33.07 ± 0.26°C). Heating rods served as the heat source, raising the temperature to the final target level before introducing the experimental fish. Each group comprised 3 parallel replicates, with 20 fish per replicate, totaling six 100 L cylindrical water tanks. During the experiment, no water circulation was employed. Heating rods were used to maintain water temperature. Dissolved oxygen levels were maintained above 6.0 mg/L, pH ranged between 7.5 and 7.8, ammonia nitrogen concentrations <0.1 mg/L, and a photoperiod of 12 hours of light followed by 12 hours of darkness was applied.
Sample collection and chemical analysis
The fish were sampled at 0, 3, 6, 12, 24, and 48 hours, with 6 fish randomly selected from each experimental group at each time point and anesthetized with MS222 (100 mg/L; Sigma Chemical Company, St. Louis, MO, USA). Then, blood was collected via the tail vein using a sterile syringe. An initial 20 μL aliquot was taken for hematological analysis. The remaining blood was centrifuged at 4 °C and 3000 rpm for 10 minutes, and the resulting supernatant was transferred to a −80 °C ultra-low temperature freezer for storage.
The ALT, AST, ALP, TP, ALB, and Glu were measured on a Beckman Coulter Cx-4 fully automated biochemical analyzer (Beckman Coulter, USA). The test kits were purchased from Nanjing Jiancheng Bioengineering Institute.
The WBC, RBC, HGB concentration and HCT were measured using the Auto Hematology Analyzer (BC-5300Vet; Mindray, P.R. China). The test kits were purchased from Shenzhen Mindray Medical Co., Ltd. (P.R. China).
Statistical analyses
All data are expressed as mean ± standard error of the mean (mean ± SEM). Statistical analysis was performed using SPSS 26.0 software (SPSS, Chicago, IL, USA). Significant differences among treatments were determined using Duncan’s multiple range test. P values < 0.05 were considered statistically significant.
Results
Plasma parameters
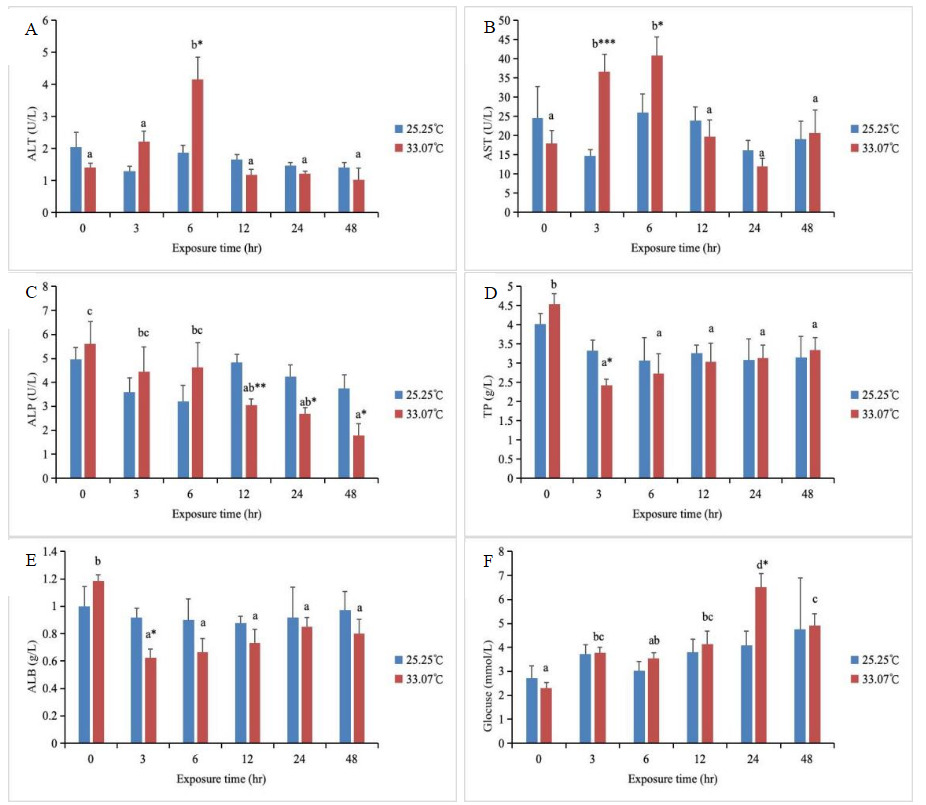
In the high-temperature treatment group, both ALT and AST activities initially increased and then decreased with increasing stress duration (Fig.1). Specifically, ALT activity peaked at 6 hours and was significantly higher than in other groups (P < 0.05), while AST activity reached its maximum at both 3 and 6 hours and was significantly higher than in other groups (P < 0.05). Additionally, at 6 hours, ALT activity in the high-temperature group was significantly higher than that in the control group (P < 0.05). At both 3 and 6 hours, AST activity in the high-temperature group was significantly higher than that in the control group (P < 0.05). Starting from 12 hours, both ALT and AST activity returned to control group levels (P > 0.05). ALP activity in the high-temperature group showed a decreasing trend with increasing stress duration, reaching its minimum at 48 hours and being significantly lower than at 0, 3, and 6 hours (P < 0.05). From 0 to 6 hours, ALP activity in the high-temperature group did not differ significantly from the control group. However, starting at 12 hours, ALP activity was significantly lower than in the control group (P < 0.05). The TP and ALB levels in the high-temperature group peaked at 0 hours and were significantly higher than those in other groups (P < 0.05). At 3 hours, both TP and ALB levels in the high-temperature group were significantly lower than those in the control group (P < 0.05). Starting from 6 hours, both TP and ALB levels recovered to the control group level (P > 0.05). The Glu content in the high-temperature group showed an initial increase followed by a decrease with increasing stress duration, reaching its peak at 24 hours and remaining significantly higher than in other groups (P < 0.05). At 24 hours, the Glu content in the high-temperature group was significantly higher than that in the control group (P < 0.05). At other time points, the Glu content in the high-temperature group did not differ significantly from that in the control group (P > 0.05). With changes in stress duration, no significant alterations were observed in the control group’s ALT, AST, and ALP activities, nor in TP, ALB, and Glu levels (P > 0.05, Fig.1).
_alt__(b)_ast__(c)_alp__(d)_tp__(e)_alb__and_(f.png)
Hematological parameters
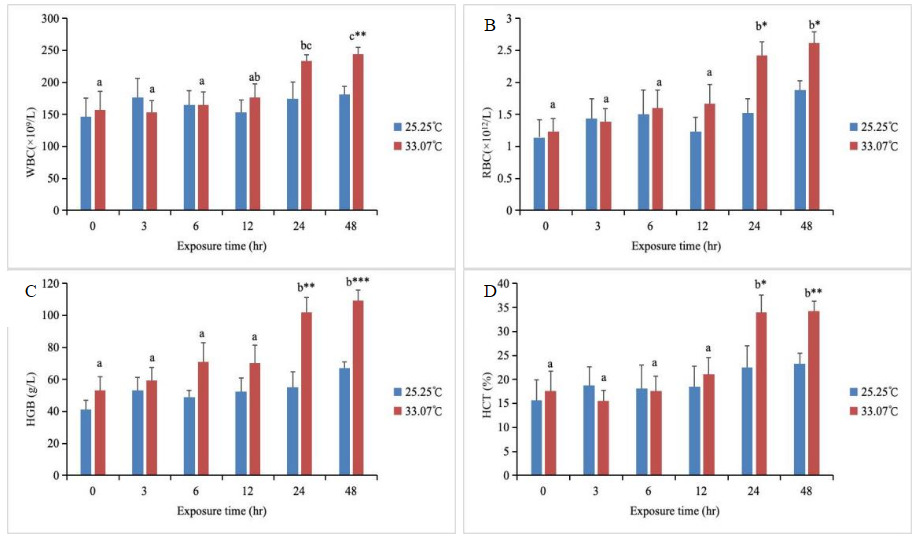
The WBC levels in the high-temperature treatment group showed an upward trend with increasing stress duration, reaching a peak at 48 hours and significantly exceeding those of other groups (except at 24 hours) (P < 0.05) (Fig. 2). Furthermore, between 0 and 24 hours, WBC levels in the high-temperature group did not differ significantly from the control group (P > 0.05), whereas at 48 hours, WBC levels in the high-temperature group were significantly higher than those in the control group (P < 0.05). RBC, HGB, and HCT levels in the high-temperature group showed an upward trend with increasing stress duration, reaching peak values at both 24h and 48h, and were significantly higher than those in other groups (P < 0.05). At 0-12 hours, the RBC, HGB, and HCT levels in the high-temperature group did not differ significantly from those in the control group (P > 0.05). However, at 24 and 48 hours, the RBC, HGB, and HCT levels in the high-temperature group were significantly higher than those in the control group (P < 0.05). Throughout the stress period, the control group exhibited no significant changes in WBC, RBC, HGB, or HCT levels (P > 0.05, Fig. 2).
_wbc__(b)_rbc__(c)_hgb_and_(d)_hct_levels_in_th.png)
Discussion
Changes in water temperature alter fish physiological functions, energy metabolism, and immune responses,19,20 thereby affecting the stability of the fish’s internal environment. Understanding the physiological responses and impacts of heat stress on key plasma biochemical parameters and hematological parameters in major aquaculture species is crucial. This study revealed that elevated temperatures significantly affected total protein, glucose, other plasma biochemical parameters, and blood cell counts in M. amblycephala under controlled conditions at 25°C and 34°C over a 48-hour period.
ALT and AST are mitochondrial transaminases. Mitochondrial bioenergetics governs environmental adaptation and stress tolerance in all poikilothermic animals, including fish. Mitochondrial function is highly sensitive to environmental temperature, as temperature directly alters physiological and biochemical rates and influences mitochondrial responses to the intracellular environment.21–23 ALT and AST are released into the bloodstream in large quantities when tissue damage or organ dysfunction occurs, leading to increased cell membrane permeability and resulting in a significant elevation in transaminase activity. These two transaminases play crucial roles in balancing amino acid levels and mediating transformations between proteins, lipids, and carbohydrates within the body.24 When environmental stressors disrupt cellular homeostasis, enzyme activity is altered accordingly. Consequently, ALT and AST serve as effective biomarkers for monitoring physiological changes in fish exposed to environmental stress.25 In this study, both ALT and AST activities in the 34°C heat-treated group exhibited a trend of initial increase followed by decrease as stress duration prolonged. ALT activity significantly increased and peaked at 6 hours, while AST activity significantly increased and peaked at both 3 and 6 hours. This indicates that liver function in blunt snout bream was impaired within 3–6 hours of heat stress exposure and began gradual recovery after 12 hours. Previous studies have demonstrated that under heat stress,18 both ALT and AST activities in blunt snout bream increase sharply at 6 hours. It is therefore inferred that the elevation of ALT and AST activities during heat stress indicates that high temperatures cause certain damage to fish organ tissues, adversely affecting their physiological functions. Therefore, the initial phase of heat stress (≤6 hours) represents a critical window for liver injury. In practical aquaculture, the first 6 hours after summer heat exposure or transportation should be considered a high-risk period. Enhanced monitoring of dissolved oxygen levels in the water is essential to alleviate the metabolic burden on the liver. ALP is a non-specific phosphohydrolase in macrophage lysosomes and an important enzyme in phosphorus metabolism within the body.8 It possesses dual defensive and digestive functions, participating in phosphate group transport and metabolism, calcium-phosphorus regulation, and other processes.26 Serum ALP activity has been identified as a component of the innate immune system in fish,27 exhibiting corresponding variations influenced by factors such as the organism’s growth stage, feeding status, and environmental conditions.28,29 In this study, as high-temperature stress persisted, ALP activity in the high-temperature group exhibited a decreasing trend. At the onset of stress, ALP activity in the high-temperature group showed no significant difference from the control group. However, starting from 12 hours, ALP activity in the high-temperature group was significantly lower than that in the control group. This indicates that heat stress may suppress macrophage function and phospho-metabolic pathways, thereby weakening the body’s immune defense capabilities. This finding is consistent with studies showing decreased ALP activity in Oncorhynchus mykiss under heat stress.30
Glu serves as a vital energy source for fish, and environmental stress can also regulate cellular energy metabolism. When fish cells are exposed to stressors, most glucose is converted into glycogen, enabling rapid energy supply.31,32 Glu is highly sensitive to changes in the body’s physiological state and to stimuli from external environmental factors, making it an important indicator in physiology, pathology, and toxicology.33 In this study, the Glu content in the high-temperature group showed an initial increase followed by a decrease with prolonged stress duration, reaching a peak at 24 hours. At this time point, the Glu content in the high-temperature group was significantly higher than that in the control group. During the early stage of the experiment, as stress duration increased, metabolic activity intensified, leading to corresponding increases in glycogen utilization and Glu synthesis rates, thereby elevating Glu levels. This indicates that fish can counteract adverse stress by elevating plasma Glu concentrations.34,35 However, as stress prolonged, Glu was extensively consumed for energy production, glycogen was broken down, and Glu levels subsequently declined. Studies on Gadus morhua36 revealed that transferring fish from a 10°C environment to 16°C and 18°C resulted in Glu content increases of 1.39-fold and 1.74-fold, respectively, compared to the control group. In contrast, research on Carassius auratus37 showed that Glu content initially increased with increasing temperature before declining, indicating a distinct energy-regulatory phase during thermal stress in fish. In aquaculture management, the proportion of easily digestible carbohydrates in feed can be appropriately increased before anticipated high temperatures or during the initial stress phase. During prolonged high temperatures, however, a continuous energy supply must be maintained to prevent metabolic failure caused by glycogen depletion. Serum contains ALB and globulin (GLO), collectively referred to as TP. Changes in TP levels directly reflect protein metabolism and synthesis within fish.38 Serum proteins are primarily synthesized in the liver, and the extent of liver damage affects protein synthesis. Serum TP and ALB serve as indicators of liver injury39; measuring their levels provides timely reflection of the liver’s protein synthesis capacity. In this study, the TP and ALB levels in the high-temperature group were significantly higher than those in other groups at 0 hours. At 3 hours, the TP and ALB levels in the high-temperature group were significantly lower than those in the control group, but subsequently recovered to the control group level. This suggests that the initial decrease in TP and ALB levels may result from impaired protein synthesis due to disrupted liver function. As stress duration increased, TP and ALB levels rose again, indicating that the liver adapts to high temperatures by upregulating relevant gene expression, significantly accelerating the synthesis rate of ALB and other plasma proteins. The pattern of TP and ALB levels, first declining and then recovering, demonstrates that the liver’s synthetic capacity is relatively unaffected by high-temperature stress and can achieve stable recovery.
Hematological parameters are commonly used to assess the physiological state of fish under stress.40,41 White blood cells serve as the body’s first line of defense and play a crucial role in the innate immune system of fish.42 In this study, WBC levels in the high-temperature group increased with increasing stress duration, peaking at 48 hours. This indicates that fish stress was most intense at 48 hours, and the changes in WBCs during stress represent a decline in the fish’s nonspecific immunity (Gholami et al., 2013). The levels of RBCs and HGB are related to oxygen transport in fish,43 with HGB also serving as the primary oxygen-carrying protein within RBCs, while simultaneously influencing the number and function of RBCs.44 In this study, the levels of RBC, HGB, and HCT in the high-temperature group increased with increasing stress duration, with significant increases observed at 24h and 48h. In hybrid yellow catfish (Pelteobagrus fulvidraco × P. vachelli),13 levels of RBC, HGB, and HCT all significantly increased under heat stress conditions. Rising water temperatures directly elevate fish metabolic rates, leading to increased RBC production and higher HGB content. The subsequent rise in HCT is a direct consequence of increased RBC numbers. Collectively, these adaptations enhance oxygen-carrying capacity to meet heightened oxygen demands.45 These regulatory mechanisms have a tolerance threshold; prolonged exposure to high temperatures may cause compensatory failure, resulting in decreased physiological indicators. This may lead to increased blood viscosity and heightened circulatory burden. Therefore, in aquaculture, regular monitoring of blood parameters is essential, particularly during high-temperature seasons. Circulatory and immune stress can be alleviated through measures such as aeration or shading. Overall, under heat stress, M. amblycephala rapidly recognizes and repairs damage, evidenced by increased ALT and AST activity and restoration of TP and ALB levels. It dynamically regulates energy via a Glu concentration pattern that first rises, then falls. Oxygen supply is ensured by the coordinated increase in RBC, HGB, and HCT levels. Immune resource allocation is adjusted through elevated WBC levels and suppressed ALP activity. Collectively, these elements form a phased, multi-system coordinated stress response program.
Conclusion
High-temperature stress dynamically affects plasma biochemical and hematological parameters over time. Biochemically, ALT and AST activities initially rose, then fell, indicating transient liver damage and repair. ALP activity decreased consistently, suggesting metabolic disruption. TP and ALB levels initially dropped but recovered, suggesting temporary inhibition of protein synthesis. Glu levels increased later, possibly due to stress-induced glucose regulation. Hematologically, WBC levels rose late, indicating immune activation. RBC, HGB, and HCT peaked at 24h and 48h, enhancing oxygen-carrying capacity. Control-group parameters remained stable, confirming the effects of heat stress. Overall, heat stress impacts liver, metabolic, and immune functions, with some recovery suggesting heat adaptation.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (32202913), the Natural Science Foundation of Shanghai (22ZR1427300), the earmarked fund for CARS-48, the China Postdoctoral Science Foundation (2024M761452), and school-level research projects of the Yancheng Institute of Technology (xjr2022043).
Authors’ Contribution
Conceptualization: Chen Chen, Wuxiao Zhang, Aqin Chen; Data curation: Chen Chen, Aimin Wang; Formal analysis: Chen Chen; Funding acquisition: Wuxiao Zhang, Silei Xia, Zhigang Yang; Investigation: Hongyan Tian; Methodology: Silei Xia, Zhigang Yang; Project administration: Wuxiao Zhang, Aqin Chen; Resources: Fei Liu; Software: Wenping Yang; Supervision: Aimin Wang; Validation: Chen Chen, Wuxiao Zhang; Visualization: Yebin Yu; Writing - original draft: Chen Chen; Writing - review & editing: Chen Chen, Wuxiao Zhang.
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
The article adheres to the Convention on Biological Diversity and the Convention on Trade in Endangered Species of Wild Fauna and Flora Research
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.