Introduction
Most aquaculture farms adopt high-density aquaculture. Owing to a lack of scientific management experience and a lack of disease prevention and control, these high-density conditions often cause problems such as frequent disease, prolonged growth cycles and inconsistent quality of economic aquatic animals. Some farmers supply chemical feed additives to increase the stress resistance and survival rate of aquatic animals. When a disease emerges, the large-scale use of antibiotics leads to problems such as high levels of drug residues in aquatic animals, increased drug resistance, and water pollution. These issues severely restrict the healthy development of the aquatic industry.
Procambarus clarkii, commonly known as crayfish, is an important economic aquaculture species in China. It is deeply loved by consumers because of its rich nutrition and delicious meat. At present, it is the most economically important freshwater crustacean in China.1 With intensive aquaculture, the yield tends to increase, as does the aquaculture utilization rate. On the one hand, density, which affects the growth of aquatic animals, also has an effect on the physiology and quality of aquatic animals. While under high-density conditions for long durations, cultured varieties are subjected to stress, which has a negative impact on physiological and biochemical functions and weakens immunity, resulting in the occurrence of diseases, particularly in crustaceans, which are protective of their domain and may exhibit fighting behaviors. Density-related factors have a significant effect on survival and growth. On the other hand, an increase in aquaculture density can lead to an increase in the water quality index of aquaculture water bodies, and the accumulation of ammonia nitrogen and nitrite nitrogen produced by aquatic animal metabolism can worsen the quality of aquaculture water bodies, resulting in large-scale disease outbreaks and death in aquaculture animals,2 such as outbreaks of white spot syndrome virus (WSSV).3 During the process of traditional shrimp culture, antibiotics are often used to treat diseases, but this leads to the production of drug-resistant pathogens and to residues of antibiotics in cultured products, which affects the quality and food safety of adult shrimp. In response to environmental protection policies, and in light of catering to consumers’ green consumption concept and improving the efficiency of aquaculture, many scholars at home and abroad have conducted abundant research on the application of Chinese herbal feed additives in aquaculture.4 Houttuynia cordata (HC) is an herb with a distinct fish-like smell; it is known as “the first natural antibiotic” and is widely distributed in China, Japan, Korea, and Southeast Asia.5 HC is known to have important drug activities, such as antitumor, antileukemic, antioxidant, and antiviral activities, and it can also be a supplement. There are still many unknown factors related to the different biological functions of the main components of HC. Although studies on humans and livestock with HC have attracted increasing attention, there is little research on the application of HC in aquaculture.
Materials and Methods
Experimental animals
Healthy crayfish were purchased from Jiangsu Jinfeng Agricultural Technology Co., Ltd., with an average weight of 6.80 ± 0.04 g. A total of 1800 crayfish were randomly allocated to eighteen pools (200 cm × 100 cm × 20 cm, polyvinyl chloride (PVC) pool). A small amount of Hydralla verticillata was included in the aquaculture pools to allow the crayfish to hide and isolate them from one another to increase their survival rate. After the crayfish were maintained under aquaculture conditions for one week, they were randomly divided into six groups (i.e., 100 crayfish·tank-1, 3 pools·group-1). The water temperature was 25.0 ± 1.0 °C, the pH ranged from 7.70–8.50, and the dissolved oxygen content was > 6.6 mg·L-1.
Feed preparation and experimental groups
The crayfish were fed basal feed, and the composition is shown in Table 1. Experimental HC was obtained from the Jiangsu Hospital of Traditional Chinese Medicine. The feed was stored at -20 °C. The experiment was divided into six groups: the basal diet group (0%), the 1.0% HC group, the 1.5% HC group, the 2.0% HC group, the 2.5% HC group, and the 3.0% HC group (Table 1, produced by Sheyang Liuhe Feed Co., Ltd.). Each group was divided into three parallel trials, in which 50 crayfish were randomly placed. During the experiment, the water temperature was 25 ± 2 °C, and the pH value was 8.0 ± 0.1. One-third of the aquaculture water was replaced by a filter screen membrane every day.
Sample collection and procedures
After the 8-week feeding experiment, the crayfish in each pool were fasted for 24 h and compressed on ice for 10 min to maintain hypothermic anesthesia. Before sampling, each individual crayfish was weighed to calculate growth performance parameters (weight gain, WG; specific growth rate, SGR; feed conversion ratio, FCR; survival rate, SR). Afterward, 3 crayfish were randomly selected from each pool to evaluate their antioxidant capacity and immune responses, and another fifty were randomly selected and used for subsequent challenge experiments. To evaluate the antioxidant capacity and immune response, hemolymph was collected from each group and mixed with anticoagulant (20 mM EDTA, pH 5.6) at a 1:1 ratio, with the continued application of ice. In addition, the hepatopancreas was rapidly frozen in liquid nitrogen and preserved at -80 °C for RNA isolation and subsequent analysis.
A whole hepatopancreas homogenate was prepared from approximately 0.3 g of hepatopancreas tissue. The crushed hepatopancreas tissue was homogenized with an Ultra-Turrax homogenizer (Tekmar Company, Cincinnati, Ohio, USA) in a 0.86% frozen normal saline solution (W/V, 1:9) and centrifuged for 10 min at 5000 r/min and 4 °C to obtain the supernatant for further analysis. The biuret method was used to determine the total protein content of the hemocyte and hepatopancreas supernatants.
Measurement of the antioxidant enzyme activities in the hemolymph and hepatopancreas
Before the enzyme activities or contents were analyzed, the aggregated hemolymph and hepatopancreas were stored at -80 °C. In accordance with the manufacturer’s guidelines, the total antioxidant capacity (T-AOC), catalase (CAT), total superoxide dismutase (T-SOD), and glutathione peroxidase (GPx) activities and reactive oxygen species (ROS) and malondialdehyde (MDA) contents were assayed using the corresponding detection kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, Jiangsu, China).
Immune-related parameters in the hemolymph and hepatopancreas
In accordance with the manufacturer’s guidelines, the lysosome (LZM), acid phosphatase (ACP), alkaline phosphatase (AKP) activities and phenoloxidase (PO) in the hemolymph and hepatopancreas activities were assayed using the corresponding detection kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, Jiangsu, China). in the hemolymph was determined using spectrophotometry according to Ashida.
Hepatopancreas immune-related gene expression measurements
The expressions of Rab5, Rab6,6 pro-phenoloxidase (proPO), crustin1,1 ferritin (fer), metallothionein7 and 18S rRNA (β-actin)8 in the hepatopancreas tissue of crayfish were analyzed via real-time PCR. The primers, which are shown in Table 2, were synthesized by Shanghai Invitrogen Corp. cDNA was generated from 500 ng of DNase-treated RNA using an ExScript RT‒PCR Kit (Takara Co., Ltd., Dalian, China), and reverse transcription PCR and real-time PCR were performed as previously described. The mRNA expression level was determined by real-time PCR (Applied Biosystems 7300 real-time PCR system, USA) using a SYBR Green I fluorescence kit. Relative Rab5, Rab6, proPO, crustin1, ferritin and metallothionein mRNA levels were quantified using the 2-ΔΔCT method, and PCR efficiency was measured using serial dilutions of cDNA according to a standard curve: ΔΔCT = (CT, target - CT, β-actin)time x - (CT, target - CT, β-actin)time 0. TRIzol reagent, RQ1 RNase-Free DNase and the ExScript RT‒PCR Kit were obtained from Dalian Takara Co., Ltd.
Crayfish culture for the challenge experiments
Crayfish with WSSV were purchased from Yellow Sea Fisheries Research, Qingdao, China. WSSV in crayfish was purified by differential centrifugation. WSSV was injected into the last abdominal segment of crayfish (2.6 × 107 virions per crayfish) for each experimental group. Mortality was recorded at 0, 24, 48, 72, 96, 120 and 168 h after WSSV infection. The challenge experiment was conducted in a separate tank.
The cumulative mortality (%) was calculated based on the following formula: cumulative mortality (%) = 100
Statistical analysis
Statistically significant differences were determined using Duncan’s multiple range test and independent sample t tests using SPSS (version 18.0) software. The data are expressed as ± SEM. The data were tested for homogeneity and subjected to an analysis of variance before the post hoc test.
Results
Effect of HC on crayfish growth
Table 3 shows the effects of HC on crayfish growth. Compared with the control diet, all five HC-supplemented diets (HC10, HC15, HC20, HC25, and HC30) produced no significant increases in final body weight (FBW), WG or SGR and no significant decreases in the feed conversion ratio (FCR) (P > 0.05). There were no significant differences in survival among the five treatment groups and the control group (P > 0.05).
Effect of HC on hemocyte and hepatopancreas antioxidant capabilities in crayfish
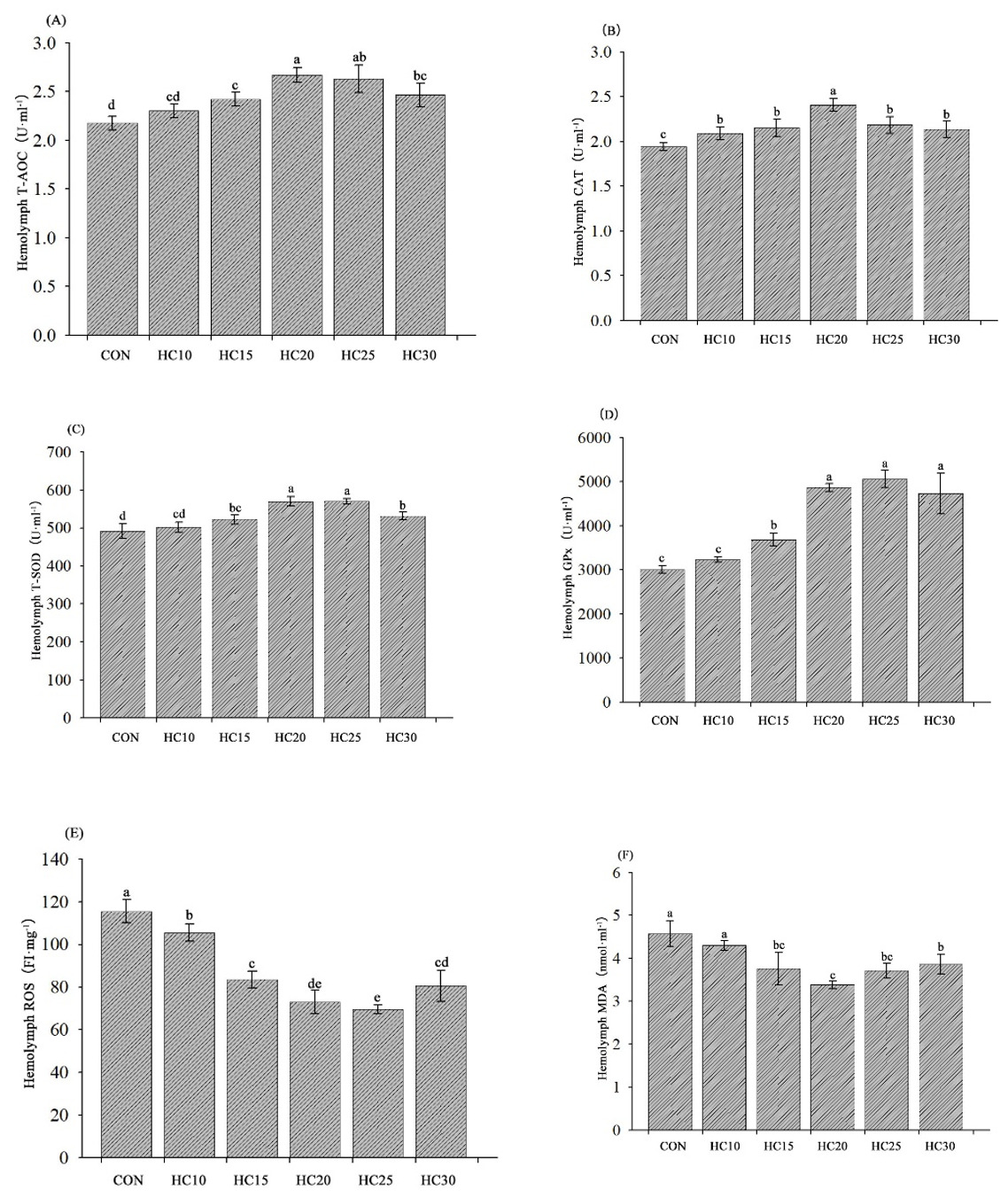
The hemocyte antioxidant capabilities of the crayfish are presented in Fig. 1. The T-AOC and T-SOD and GPx activities were significantly greater and the MDA content was lower in the hemolymph from the crayfish fed the HC15, HC20, HC25, and HC30 diets than in the hemolymph from the crayfish fed the control diet (P < 0.05) (Fig. 1A, C, D and F). Compared with the control diet, all five HC-supplemented diets (HC10, HC15, HC20, HC25 and HC30) significantly increased CAT activity and decreased the ROS content (P < 0.05) (Fig. 1B and E).
__cat(b)__t-sod_(c)__gpx_(d)_and_the_contents_of_ros_(e)_and_md.png)
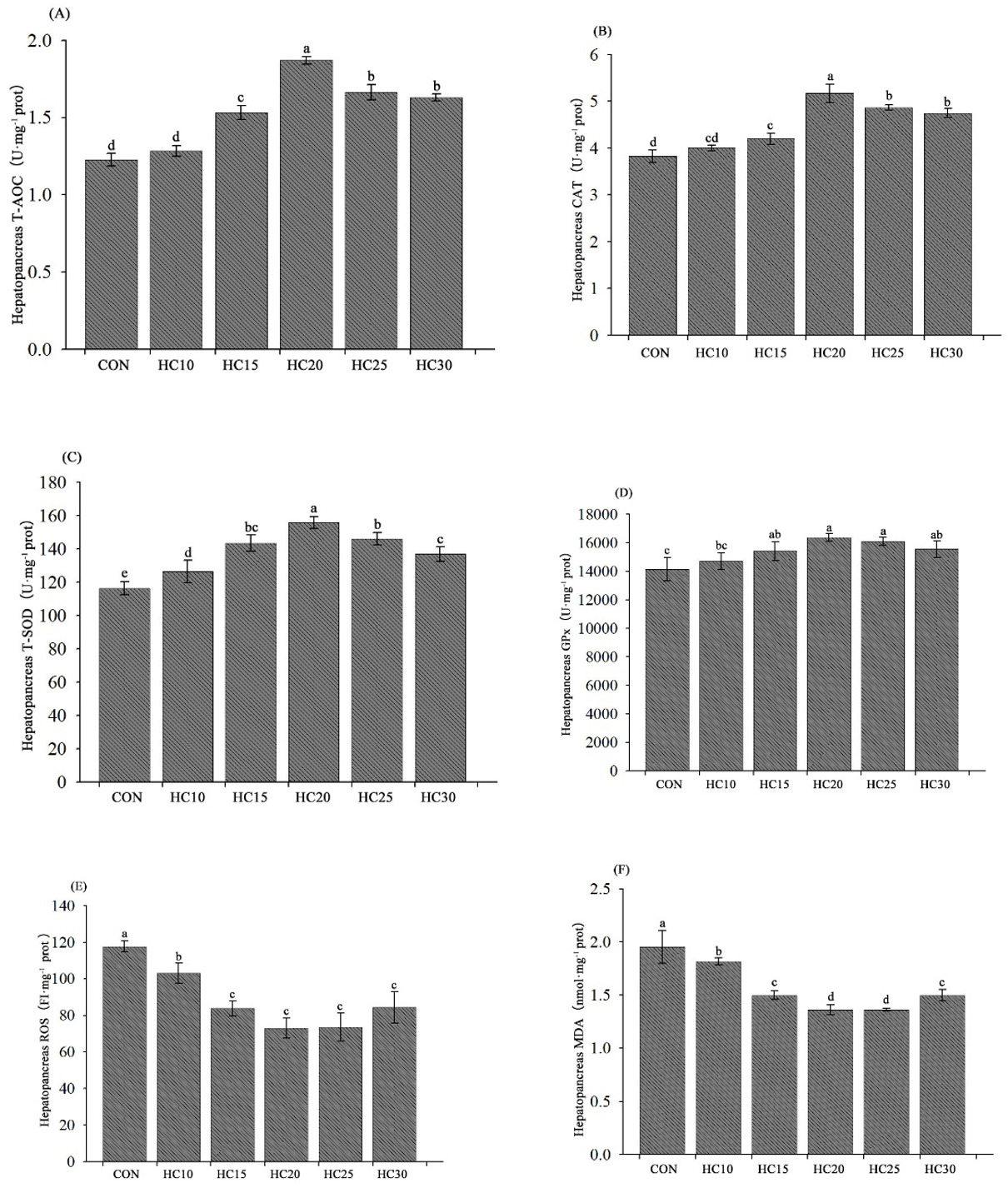
The hepatopancreas antioxidant abilities of the crayfish are presented in Fig. 2. The T-AOC and CAT and GPx activities were significantly greater in the hemolymph from the crayfish fed the HC15, HC20, HC25, and HC30 diets than in the hemolymph from the crayfish fed the control diet (P < 0.05) (Fig. 2A, B and D). All five HC-supplemented diets (HC10, HC15, HC20, HC25 and HC30) significantly increased T-SOD activity (Fig. 2C) and decreased the MDA and ROS contents (Fig. 2E and F).
__cat(b)__t-sod_(c)__gpx_(d)_and_the_contents_of_ros_(e)_and_md.png)
Effects of HC on hemocyte and hepatopancreas immune-related parameters in crayfish
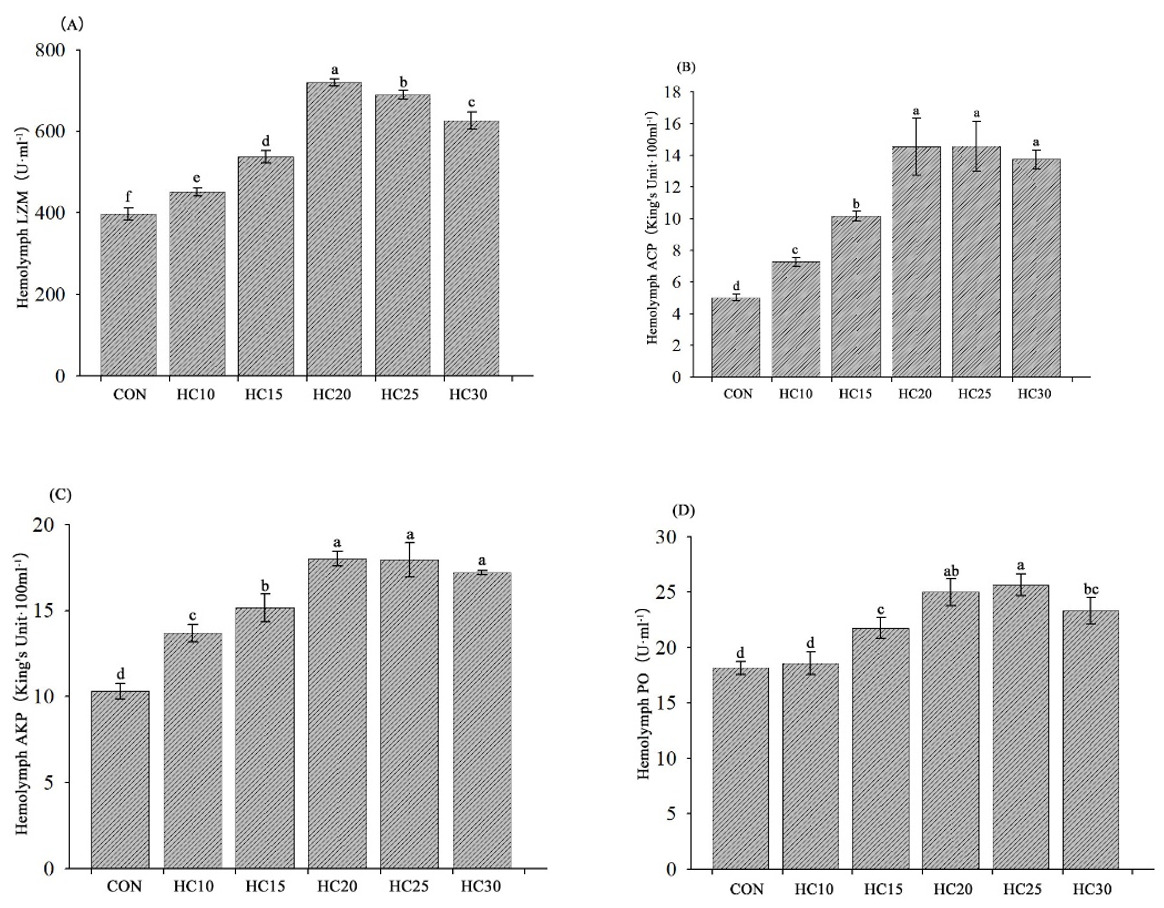
The immune-related parameters of the crayfish are presented in Fig. 3. Compared with the control diet, the five HC-supplemented diets (HC10, HC15, HC20, HC25 and HC30) significantly increased LZM, ACP and AKP activities (P < 0.05) (Fig. 3A, B and C). The hemocyte PO content was greater in the crayfish fed the HC15, HC20, HC25, and HC30 diets than in the crayfish fed the control diet (P < 0.05) (Fig. 3D).
__acp_(b)__akp_(c)_and_po_(d)_in_crayfish_hemolymph_subjected_to_di.png)
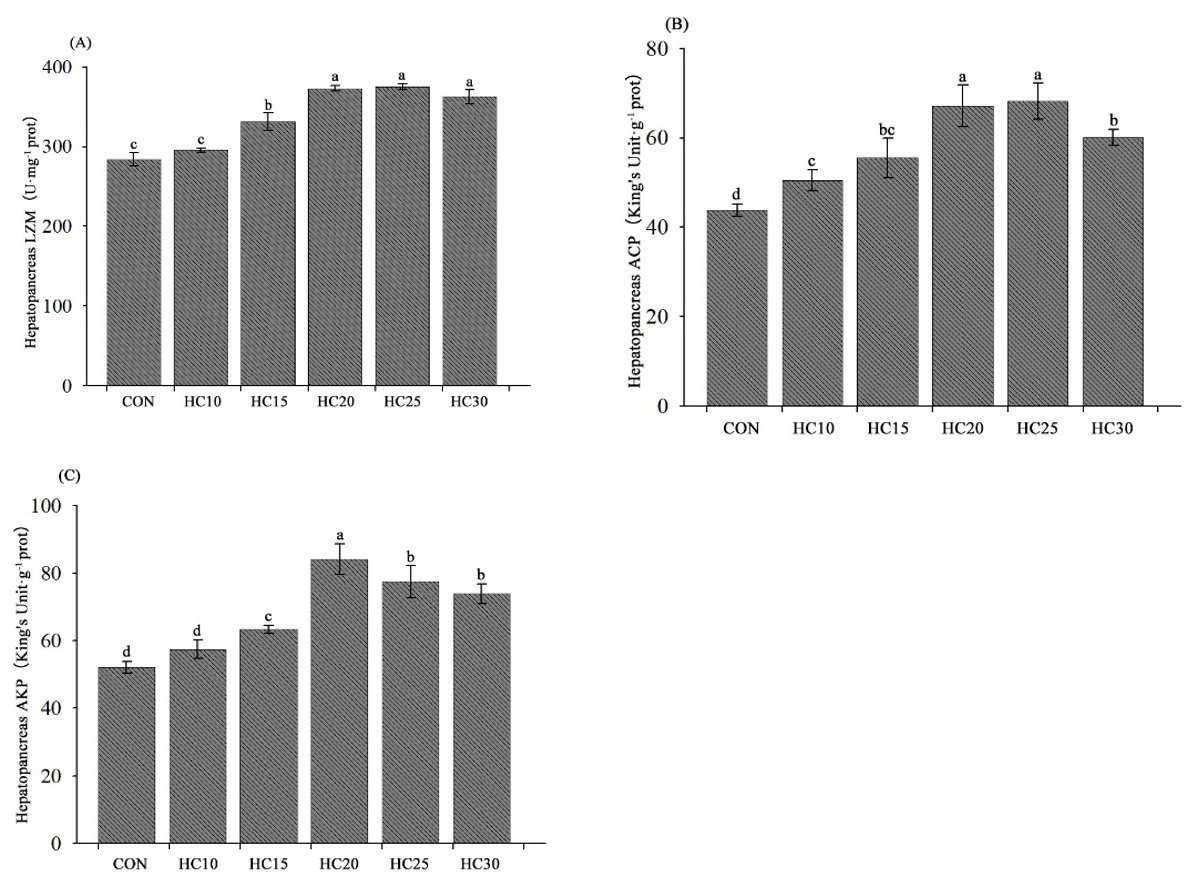
The hepatopancreas immune-related parameters of the crayfish are presented in Fig. 4. LZM and AKP activities were significantly greater in the hemolymph from the crayfish fed the HC15, HC20, HC25, and HC30 diets than in the hemolymph from the crayfish fed the control diet (P < 0.05) (Fig. 4A and C). Compared with the control diet, all five HC-supplemented diets (HC10, HC15, HC20, HC25 and HC30) significantly increased ACP activity (P < 0.05) (Fig. 4B).
__acp_(b)_and_akp_(c)_in_crayfish_hepatopancreas_subjected_to_diffe.png)
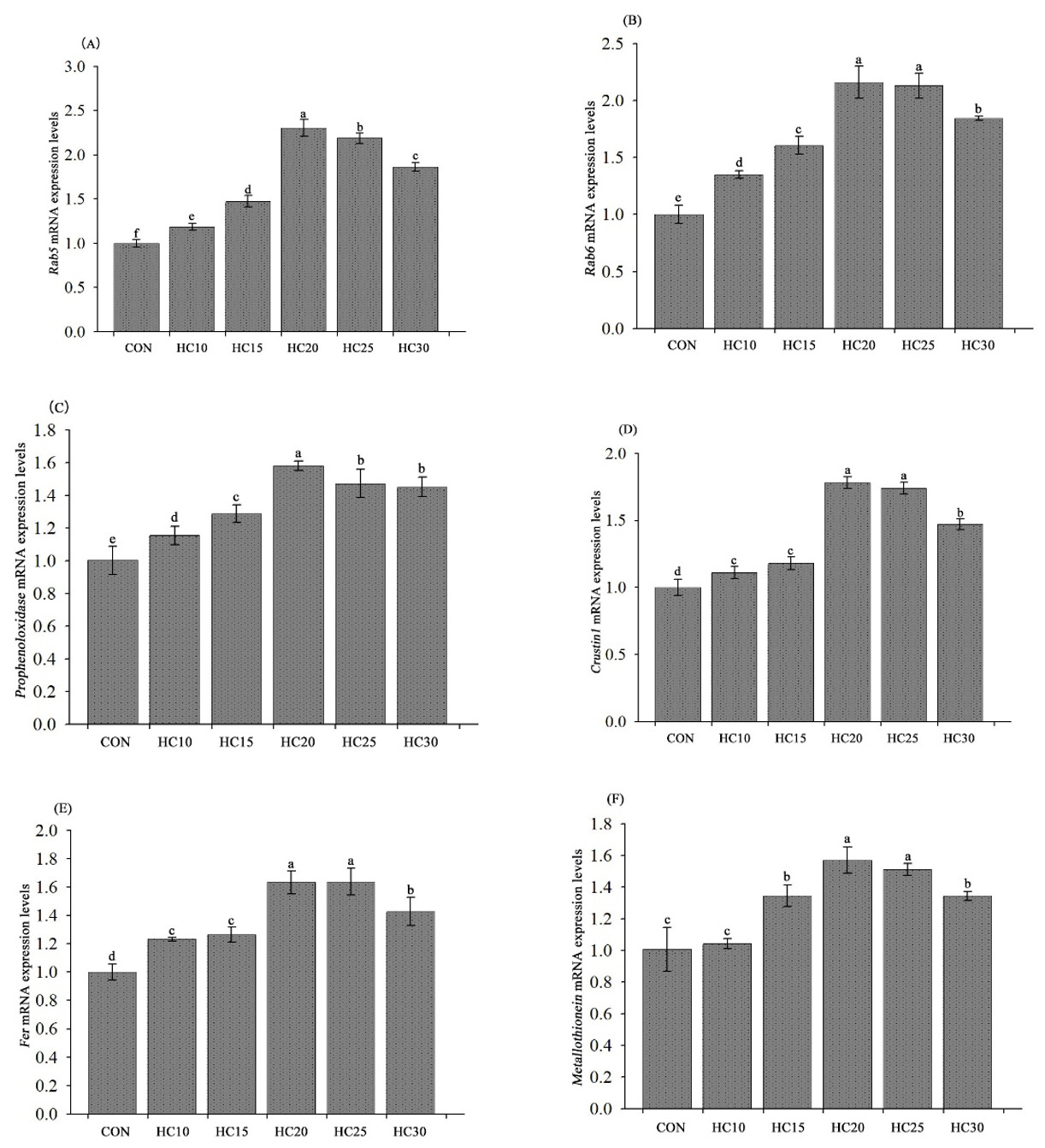
Effects of HC on the expression of genes related to antioxidants and immunity in the crayfish hepatopancreas
Compared with the control diet, all five HC-supplemented diets (HC10, HC15, HC20, HC25 and HC30) produced significantly increased hepatopancreas-relative Rab5, Rab6, proPO, crustin1 and ferritin mRNA levels (P < 0.05) (Fig. 5, A, B, C, D and E). The hepatopancreas-relative metallothionein mRNA expression levels were significantly greater in the hemolymph from the crayfish fed the HC15, HC20, HC25, and HC30 diets than in the hemolymph from the crayfish fed the control diet (P < 0.05) (Fig. 5F).
_and_rab6_(b)__propo_(c)__crustin_1_(d)__ferritin_(e)_and_.png)
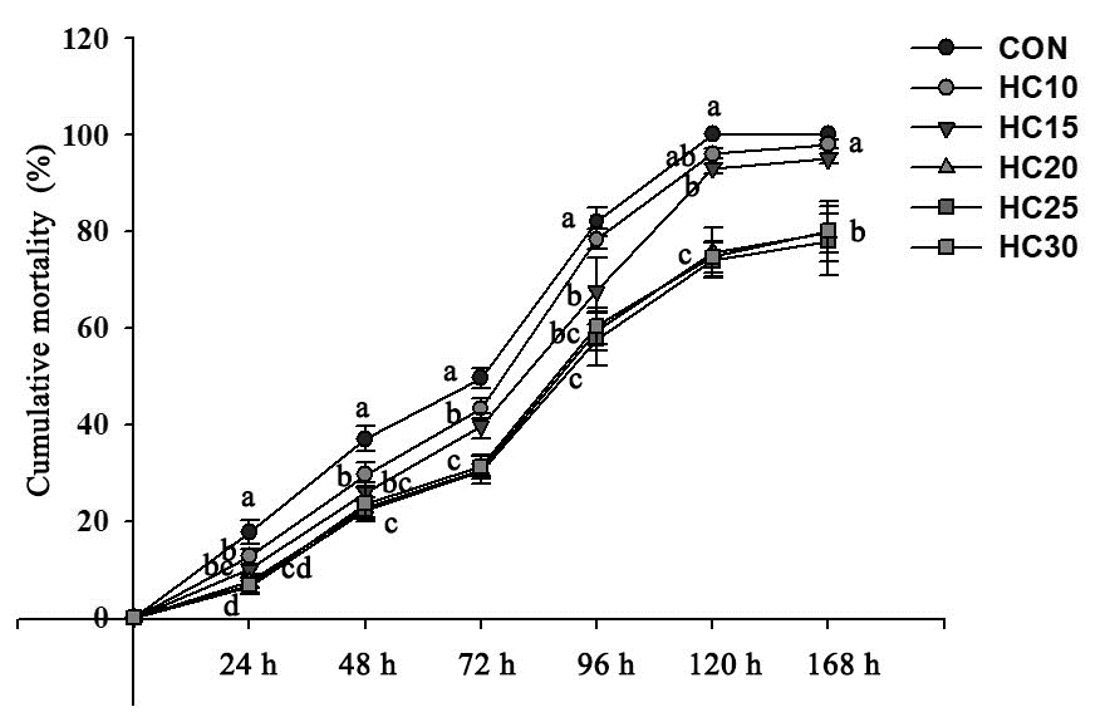
Effect of HC on the crayfish challenge test results
The cumulative mortality rate after WSSV infection is shown in Fig. 6. The greatest increase in the cumulative mortality rate occurred 48 to 120 h after infection, and the total cumulative mortality rates at the end of the experiment were 100.00%, 98.00%, 95.00%, 79.67%, 78.00% and 80.00% for the control diet and five HC-supplemented diets (HC10, HC15, HC20, HC25 and HC30), respectively. The cumulative mortality rates were significantly greater at 72, 96, 120 and 168 h after infection in the control crayfish and in the crayfish fed the HC10 diet than in the crayfish fed the HC15, HC20, HC25 and HC30 diets (P < 0.05, Fig. 6).
_infec.png)
Discussion
Effect of HC on crayfish growth
Chinese herbal medicine feed additives promote the growth performance of aquatic animals by improving digestive enzyme activities and the intestinal microbial community structure.9 There are relatively few studies on HC as a feed additive for aquatic animals. Studies have shown that adding 2% HC to feed can promote the growth of juvenile grass carp (Ctenopharyngodon idellus).10 However, some studies have shown that HC has no effect on the growth of Cobia (Rachycentron canadum)11 or Nile tilapia (Oreochromis niloticus).5 All five HC-supplemented diets did not significantly increase the growth of the crayfish (P < 0.05). However, the diets supplemented with HC significantly reduced the mortality rate after infection with WSSV in this study, which is consistent with results from studies of glycyrrhizic acid,9 hesperidin,12 and Astragalus polysaccharides.13 Therefore, as a natural antibiotic, HC plays an important role in resisting WSSV. Furthermore, the main reasons for the improved growth in juvenile grass carp (Ctenopharyngodon idellus) may be improved intestinal flora structure and function. We also speculate that the reason for the lack of improvement in the growth of crayfish may be the poor palatability of HC, which is not conducive to the attractiveness of the food.
Effects of HC on the antioxidant capabilities of crayfish
When crustaceans are under emergency stress, such as environmental stress and disease-induced stress, the accumulation of ROS exceeds the body’s self-regulation ability and damages the body’s physiological and immune functions. Lipid peroxidation is the result of the oxidation of polyunsaturated fatty acids in the lipid membrane. As one of the products of lipid peroxidation, MDA can aggravate cell membrane damage. Therefore, the MDA content reflects the degree of ROS damage in tissue cells and indirectly reflects the antioxidant capacity of the body.14 To remove toxic ROS, aquatic animals have their own effective antioxidant defense system to protect tissues from oxidative damage. Antioxidant enzymes and proteins play important roles in resolving stress and neutralizing excess ROS. The T-AOC is a comprehensive index that has been recently applied to measure the function of the antioxidant system. It reflects the compensatory ability of antioxidant enzymes and nonenzymatic systems to detoxify external stimuli and promote the health of the body.15 The SOD system provides the first line of defense against oxidative stress by converting O2•- to H2O2 to prevent direct damage induced by O2•- to the body.16 The toxic compound H2O2 is further eliminated by GPx and CAT, which catalyze the conversion of H2O2 to H2O.17 Therefore, the three enzymes SOD, CAT and Gpx cooperate with one another to inhibit lipid oxidation in the cell membrane and reduce oxidative damage.
Studies have shown that hesperidin from dried tangerine peel can increase the cellular antioxidant capacity and enhance the antioxidant status in broiler chicks by increasing T-AOC and GPx activities and reducing the MDA content.12 Other studies have shown that astaxanthin,18 chitosan oligosaccharide19 and Astragalus polysaccharide13 can increase the SOD activity of crayfish. Kelp powder can also increase SOD and GPx activities and decrease the MDA content of crayfish, and glycyrrhizic acid can increase T-AOC and T-SOD and GPx activities and decrease the MDA and SOD contents of crayfish.9 Other studies have shown that icariin can alleviate oxidative stress caused by duck hepatitis A virus by increasing SOD and GPx activities and reducing the MDA content in ducks20; furthermore, icariin can significantly improve cisplatin-induced oxidative stress by reducing MDA and ROS levels in HEK-293 cells in vitro.21 Accordingly, we speculate that feeding crayfish an appropriate amount of HC improved the CAT and GPX activities in the hemolymph and hepatopancreas, which improved the ability to decompose residual H2O2 via increased SOD activity. Thus, the T-AOC ultimately improved, and the contents of ROS and MDA in the crayfish decreased. These findings may help us understand the physiological mechanism of the effects of HC supplementation in crayfish diets.
Effects of HC on immune-related parameters in crayfish
Lysozyme is an important nonspecific immune factor in animals and has a strong killing effect on gram-positive bacteria. The activity and antibacterial abilities of lysozyme in animals are directly related to the immune function and health of the fish. ACP is an enzyme similar to lysozyme that can catalyze the hydrolysis of phosphate monoesters. Previous studies have shown that ACP release is accompanied by an immune response, specifically crustacean blood cell phagocytosis and engulfment. Research has shown that the immunity of aquatic animals increases with increasing lysozyme and ACP activities.22 AKP is an important metabolic regulatory enzyme in organisms. It can directly participate in phosphate group transfer and calcium and phosphorus metabolism. AKP plays an important role in the absorption and utilization of nutrients by fish and shrimp and improves their disease resistance.23 Studies have shown that fructooligosaccharides24 can increase the LZM, ACP, and AKP activities in Litopenaeus vannamei, that Astragalus polysaccharide can increase the LZM activity of crayfish,13 that kelp powder can increase the AKP activity of crayfish, and that astaxanthin18 and chitosan oligosaccharide19 can increase the AKP and ACP activities of crayfish. In addition, studies have shown that HC can increase the AKP, ACP and LZM activities of sea cucumber (Apostichopus japonicus) and increase the LZM activity of juvenile grass carp (Ctenopharyngodon idellus).10
PO, one of the key immune components in the invertebrate innate immune system, usually does not have bactericidal or antibacterial activity but can catalyze the conversion of phenolic substrates to quinones, which are ultimately converted to melanin through a series of nonenzymatic reactions. Quinones have been reported to have bactericidal and bacteriostatic effects, while melanin has also been found to agglutinate pathogens, repair damage and mediate phagocytosis.25 Studies have shown that Uncaria tomentosa26 can increase PO and AKP activities in Pacific white shrimp (Litopenaeus vannamei), that glycyrrhizic acid can improve PO activity in crayfish.9 In addition, studies have shown that a floating bed of HC can increase PO activity in aquacultured genetically improved farmed tilapia (GIFT) (Oreochromis niloticus).27
Therefore, we speculate that Houttuynia cordata, as a feed additive used in Chinese herbal medicine, can improve the phagocytosis and engulfment of blood cells and enhance bactericidal ability in vivo by increasing the activities of lysozyme and ACP in crayfish. Moreover, it can catalyze the conversion of its phenolic substrate to quinone species and improve immunity. In addition, it can improve the activity of AKP in crayfish, promote nutrient absorption and increase disease resistance. Thus, LZM, ACP, and AKP activities were significantly increased in the hemolymph and hepatopancreas, and PO activity was increased in the hepatopancreas of crayfish fed the HC15, HC20, HC25 and HC30 diets, which indicates that dietary HC could enhance immunity in crayfish.
Effects of HC on the expression of genes related to immune function in crayfish
Rab proteins represent the largest branch of the Ras-like small GTPase superfamily. To date, more than 70 Rab genes have been successfully identified in the human genome. Most Rab proteins play important roles in regulating membrane transport at different levels in all eukaryotic cells. Rab5 and Rab6 are important members of the Rab family and play regulatory roles in the interaction between phagosomes and other intracellular compartments. These two genes are generally expressed in various tissues, and changes in Rab5 (or Rab6) expression are related to various diseases.28 Recent studies have shown that Rab5 and Rab6 are involved in the innate immune response of aquatic animals. Studies have shown that Rab5 mRNA expression levels are significantly increased in large yellow croaker (Larimichthys crocea) spleens after infection with Vibrio parahaemolyticus29 Rab6 is reportedly involved in the regulation of antibacterial and antiviral phagocytosis by Marsupenaeus japonicus blood cells. The phagocytic percentage and phagocytic index of blood cells have been found to be positively correlated with the expression of Rab6.30 In addition, the expression levels of the Rab5 and Rab6 genes in the crayfish hepatopancreas, gills, and intestines are significantly increased after Aeromonas hydrophila and WSSV infection, respectively.6 These results indicate that Rab5 and Rab6 play important roles in the immune response to viruses and bacteria. It is speculated that Houttuynia cordata can promote lysozyme and ACP enzyme activities in crayfish blood cells, the hepatopancreas and other tissues by increasing the gene expression of Rab5 and Rab6, resulting in high phagocytosis and bactericidal ability. Therefore, increased Rab5 and Rab6 mRNA levels could enhance crayfish immunity.
The proPO activation system involves a variety of enzyme cascade reactions. When bacteria or fungi infect shrimp, the system is activated, and proPO is activated into the active PO. Studies have shown that proPO in shrimp exists mainly in inactive GCs, which are released through exocytosis and then transformed into PO.31 Crustin 1 is an invertebrate antimicrobial protein that widely exists in shrimp, crabs, lobsters and other crustaceans. Studies have shown that proPO and crustin 1 mRNA expression levels are significantly increased in crayfish after infection with WSSV.1 These results indicate that the increase in PO activity in this study was closely related to the high expression of the proPO gene. Houttuynia cordata may increase the activity of PO by increasing the expression of the proPo gene and inducing exocytosis in GCs. Ferritin is a cellular antioxidant that regulates the availability of iron ions for the Fenton reaction to control the production of ROS.32 Metallothioneins act as antioxidants by scavenging free radicals. Studies have shown that ferritin and metallothionein mRNA expression levels are significantly increased in crayfish after exposure to nitrite and sulfide stress.7 Therefore, the high expression of the ferritin and metallothionein genes in this study was closely related to the decrease in the ROS content in the hepatopancreas. Houttuynia cordata may improve the ability of free radical scavenging by increasing the expression of the ferritin and metallothionein genes to control the production of ROS and ultimately reduce the content of ROS. Thus, as a result of the upregulation of proPO, crustin 1, ferritin and metallothionein expression, the immunity of crayfish may be increased.
In summary, the upregulated expression of Rab5, Rab6, proPO, crustin 1, ferritin and metallothionein in crayfish could lead to increased immunity in crayfish due to feeding with the HC15, HC20, HC25 and HC30 diets. However, the mechanism of this effect requires further study.
In conclusion, HC has antitumor, antileukemia, antioxidant, antiviral and other biological activities. In this study, dietary supplementation with HC significantly improved the antioxidant activity, immunity and disease resistance of crayfish. Therefore, HC is an important immunopotentiator and will be used in crayfish aquaculture in the future. Under our research conditions, the optimal dose of dietary HC was present in the HC15, HC20, HC25, and HC30 diets (1.5%-3.0%, w/w). However, further studies in different animal and cell models are necessary to comprehensively evaluate the immunopotentiation and antioxidant activity of HC and the exact molecular mechanisms involved.
Acknowledgments
This project is funded by the industry university research cooperation project of Yancheng Institute of Technology (20241129152604)
Authors’ Contribution
Conceptualization: Fei Liu (Equal), Bo-Xing Cheng (Equal). Methodology: Fei Liu (Lead). Writing – original draft: Fei Liu (Lead). Funding acquisition: Fei Liu (Equal), Ai-Ming Wang (Equal). Investigation: Chao Feng (Equal), Jian-Qiao Zhu (Equal), Meng-Xue Li (Equal), Xing-Guo Liu (Equal), Hao Zhu (Equal). Formal Analysis: Ye-Bing Yu (Equal), Wen-Ping Yang (Equal), Hong-Yan Tian (Equal). Supervision: Wen-Ping Yang (Lead). Resources: Ming Xie (Lead). Writing – review & editing: Ai-Ming Wang (Lead).
Competing of Interest – COPE
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our work, there is no professional or other personal interest of any nature or kind in any product, service and/or company that could be construed as influencing the position presented in, or the review of, the manuscript entitled, “Effects of the medicinal herb Houttuynia cordata on growth performance, antioxidant capacity, immune response and immune-related gene expression in the red swamp crayfish Procambarus clarkii”.
Ethical Conduct Approval – IACUC
All handling of crayfish was conducted in accordance with the guidelines on the care and use of animals for scientific purposes established by the Institutional Animal Care and Use Committee (IACUC) of Yancheng Institute of Technology, China. The IACUC specifically approved this study within the project “The artificial culture of red swamp crayfish (Procambarus clarkii)” (approval number YCIT (F)-18-010).
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.