Introduction
Redtail botia Yasuhikotakia modesta is a freshwater species native to the Mekong and Chao Phraya River basins and has recently gained commercial value beyond its traditional role as an ornamental fish.1–3 The species’ high market price and increasing demand have intensified interest in developing hatchery-based seed production as wild stocks continue to decline.4 Although artificial breeding techniques for redtail botia have already been established, successful larval rearing remains a significant challenge.5
Like many freshwater fish larvae, redtail botia initially rely on live feed organisms such as copepods, rotifers, Moina, and other zooplankton.4–6 These organisms provide not only essential nutrients (e.g., proteins, vitamins, carbohydrates, minerals, amino acids, and fatty acids) but also digestive enzymes that stimulate feeding, digestion, and gut development.7,8 Certain zooplankton are considered highly nutritious prey, supplying high levels of digestive enzymes that enhance larval appetite.9 In addition, the motility of live feed organisms offers sensory cues that trigger feeding behavior in fish.7,10,11 Because of their limited mouth size and swimming ability, fish larvae show a preference for live feed, which is appropriately sized and easier to capture, thus improving growth rates.12,13 This phenomenon is also observed in redtail botia, which started to capture small rotifers as early as three days after hatching (DAH), coinciding with mouth opening and the onset of exogenous feeding.4
Reliance solely on live feed poses a major challenge in fry rearing due to high costs, labor intensity, space, and infrastructure requirements, despite its nutritional advantages.14–18 In addition, reliance on live feed poses risks, including inconsistent nutritional composition and potential contamination by pathogenic bacteria.15,19,20 Artificially formulated feed (microdiets) offers a more consistent and economical alternative, but many fish species show poor survival and growth when weaned too early, largely because of immature digestive systems and insufficient digestive enzyme activity.17,21–23 Therefore, co-feeding live feed and formulated feed has been shown to be an effective transitional strategy, improving survival and growth performance while gradually stimulating digestive enzyme activity and facilitating adaptation to formulated diets.10,24,25 This approach reduces dependence on live feed, minimizes production costs, and enhances the success of larval rearing in aquaculture.10,18,25,26 However, there is no universal weaning strategy and optimal weaning protocols are species-specific.18,27 Although studies have established weaning protocols for related species such as butter catfish Ompok bimaculatus,28 angelfish Pterophyllum scalare29 and severum Heros severus,29 Chinese longsnout catfish Leiocassis longirostris Günther,30 Clarias magur,31 and climbing perch Anabas testudineus,32 no study has systematically evaluated weaning strategies or determined the optimal weaning time for redtail botia Yasuhikotakia modesta. In particular, the relationship between weaning timing and the development of digestive enzyme activity in this species remains insufficiently understood. This lack of information represents a critical bottleneck for the development of efficient hatchery protocols.
Therefore, the present study evaluated the effects of weaning redtail botia larvae from Moina to a formulated feed at different post-hatching ages. The experiment assessed how the timing of weaning influences larval survival, growth performance, digestive enzyme activity (i.e., trypsin, pepsin, lipase, and amylase), and nutritional body composition. The study aims to identify the appropriate timing for weaning formulated feed that supports the efficient larval rearing of this species.
Methodology
Animal source and husbandry
The broodstock used in the experiment had an average weight of 35–50 g/fish. To stimulate artificial reproduction, the fish were injected with human chorionic gonadotropin (hCG) and luteinizing hormone-releasing hormone analogue (LHRH-a) combined with domperidone (DOM). Female fish were injected with two doses: a preliminary dose of 1,000-1,500 IU/kg and a decisive dose of 80-120 µg LHRH-a and 10 mg DOM/kg, 6 h apart. Male fish were injected with a dose equal to half the decisive dose of the female fish, and simultaneously with the decisive dose of the female fish. The female fish ovulated 6-8 h after the stimulation injection. Eggs were brushed and artificially inseminated. The eggs hatched at 29–30°C after 10–11 h.
At 2 DAH, fish larvae were reared at a density of 5 individuals per liter in a 3 m³ nursing tank supplied with diverse natural food organisms (e.g., rotifers, cladocerans, and copepods) under a temperature range of 27–30°C. At 11 DAH, 4,800 larvae were randomly transferred to experimental tanks. The experiment was conducted in plastic tanks (60 × 40 × 40 cm; water volume 40 L), each equipped with eight PVC tubes (diameter 2.7 cm; length 5 cm) that served as shelters. Each tank contained 200 larvae, corresponding to a stocking density of 5 fish per liter. The average initial larval weight was 0.0196 g, and the mean length was 1.06 cm, based on measurements from a random sample of 60 larvae taken from the same spawn that was not used in the experiment.
Experimental design
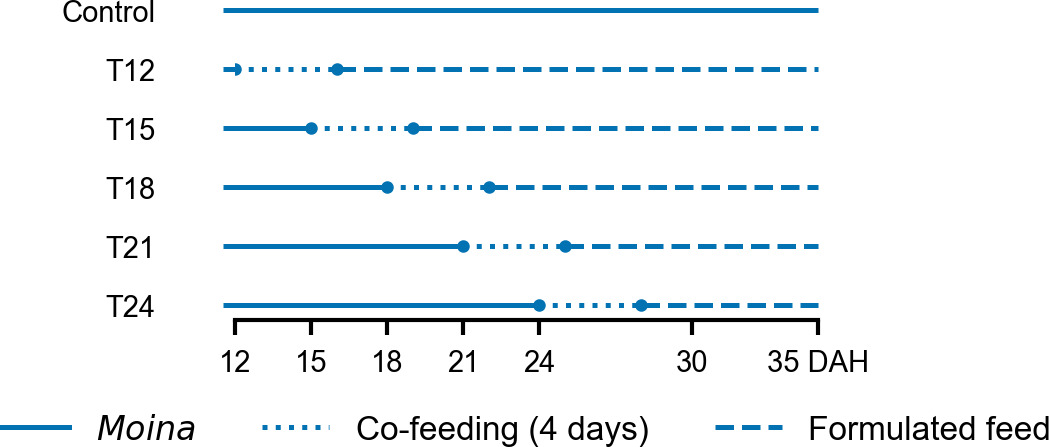
The experiment was designed with six treatments in which redtail botia larvae were weaned onto formulated feed (microdiet) starting at different DAHs: 12 DAH (T12), 15 DAH (T15), 18 DAH (T18), 21 DAH (T21), and 24 DAH (T24), along with a control group (Control) fed exclusively on Moina (Figure 1). The earliest weaning age was set at 12 DAH when gastric glands appeared, and pepsin activity emerged, indicating the functional development of the digestive system necessary for the utilization of formulated feed.4 Treatments were arranged in a completely randomized design, with each treatment replicated across four tanks. Prior to the weaning period, larvae were fed only Moina, twice daily at 08:00 and 15:00 from 3 DAH. In the formulated weaning treatments, larvae were co-fed with Moina and microdiet for a transitional period of four days, during which the proportion of microdiet was increased daily by 25% while the proportion of Moina was decreased by the same amount: (i) Day 1: 25% microdiet and 75% Moina; (ii) Day 2: 50% microdiet and 50% Moina; (iii) Day 3: 75% microdiet and 25% Moina; (iv) Day 4: 100% microdiet. The experiment ended at 35 DAH.
_of_redtail_botia_larvae_weaned_onto_microdi.png)
The nutritional composition of Moina and the microdiet is presented in Table 1. The microdiet used in this study was a commercial feed composed primarily of fish meal, lecithin, wheat gluten, dried seaweed, fish oil, maize starch, vitamins, and minerals, with a composition of 46.90% protein and 6.23% lipid. The particle size of the microdiet is 0.2–0.3 mm. Larvae were fed microdiet four times daily (06:00, 10:00, 14:00, and 16:00). Prior to feeding, the microdiet was blended with water and placed on feeding screens. Fish were fed ad libitum, with a daily feeding rate averaging approximately 20%. Feeding screens were cleaned and tank bottoms were siphoned daily to remove feces. Water exchange (40–60%) was performed each morning before feeding. Water quality was monitored daily, with temperature (HI98127, Hanna Instruments, USA) ranging from 27.0–28.5°C, pH (HI98127, Hanna Instruments, USA) from 7.0–7.2, and dissolved oxygen (HI9147, Hanna Instruments, USA) from 6.2–6.8 mg/L. Total ammonia concentration was analyzed weekly (HI83303, Hanna Instruments, USA) and remained below 0.5 mg/L.
Sampling and analysis
Survival and growth
At the end of the experiment, the final number of fish was counted, and the batch was weighed to determine survival and growth. Sixty fish were randomly selected from each tank to analyze the variation in length and weight, with individual body length (mm) measured by a ruler and individual body weight (g) determined using an electronic scale. During the experiment, dead fish were counted daily to assess mortality. The following parameters were calculated to assess survival and growth:
Survival rate (%) = 100 × final number of fish/initial number of fish
Specific growth rate in weight (SGR-W, %/day) = 100 × [ln(Wf)/ln(Wi)]/days of feeding Specific growth rate in length (SGR-L, %/day) = 100 × [ln(Lf)/ln(Li)]/days of feeding
where Wf/Lf and Wi/Li represent the final and initial mean body weights (g) and lengths (mm), respectively.
Coefficient of variation for weight (CV-W, %) = 100 × (standard deviation of weight/mean weight)
Coefficient of variation for length (CV-L, %) = 100 × (standard deviation of length/mean length)
Digestive enzyme activities
At the end of the experiment, two fish were collected from each tank for digestive enzyme analysis. Samples were washed with distilled water, placed in microtubes, and stored at –80°C until analysis. Enzyme assays focused on the activities of trypsin, pepsin, lipase, and amylase. Larvae samples were ice-chilled and homogenized in a buffer of 20 mM Tris-HCl, 1 mM EDTA, 10 mM CaCl2, pH 7.5.33 Digestive enzyme activities were based on the protein concentration in the sample. The Bradford method using CBB solution (Nacalai Tesque, Inc., Kyoto, Japan) was applied to determine the protein content of samples.34 Enzyme activity analysis was conducted following the procedures described in our prior study Khoa et al., 201935 including Bolasina et al., 200633 for trypsin, Worthington et al., 199336 and Rønnestad et al., 201313 for pepsin, Roberts, 198537 for lipase, and Murashita et al., 201838 for amylase.
Nutritional composition of fish
At the end of the experiment, fish from each tank were collected and stored at -20°C for analysis of proximate carcass composition. Proximate composition of the experimental diets and the initial and final body composition of fish were estimated. Moisture, crude protein, crude lipid, and crude ash were determined following standard methods.39
Moisture content was determined by weighing samples in pre-dried porcelain crucibles, followed by desiccation in a drying oven (Gallenkamp, UK) at 105°C for over 4 h (dry samples) or 24 h (wet samples) until a constant weight was achieved. Loss on drying was recorded as the moisture content. The dried samples were then incinerated in a muffle furnace (Gallenkamp, UK) at 600°C for 6–8 h, and the loss after incineration was taken as the total ash. The remaining residue after ash subtraction was considered fiber.
Crude lipid was extracted with petroleum ether in a Soxhlet apparatus and weighed after evaporation of the petroleum ether. Crude protein was determined by the Kjeldahl method (N × 6.25) using three steps: digestion with concentrated H2SO4 98% and H2O2 catalyst at high temperature in a Kjeldahl digestion system (Gerhardt, Germany), distillation after reaction with NaOH 40%, with released NH3 absorbed by H3BO3 2% in a Kjeldahl distillation system (Vapodest 10sn, Gerhardt, Germany), and manual titration with H2SO4 0.01N (Merck, Germany).
Statistical analysis
Statistical analyses were conducted using R (version 4.1.1). Results are presented as mean ± standard deviation (SD). Normality and homogeneity of variance were assessed using the Kolmogorov-Smirnov and Levene’s tests. One-way analysis of variance (ANOVA) was applied to determine significant differences. Duncan’s multiple range test was used for post-hoc comparisons. Statistical significance was set at α = 0.05.
Results
Survival rates
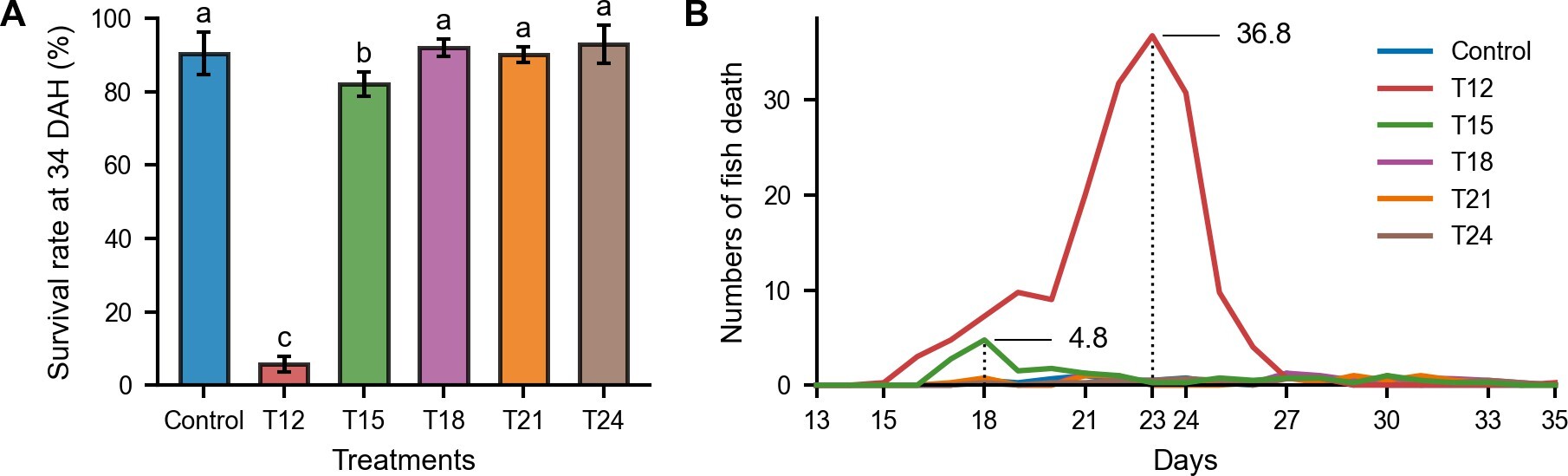
The survival rate increased with later weaning times (p<0.05; see Figure 2A). The Control group exhibited a survival rate of 90.38 ± 5.79%. T12 had the lowest survival rate (5.75 ± 2.10%), which was significantly lower than all other treatments. T15 had a higher survival rate compared to T12 (82.13 ± 3.30%), but it was significantly lower than those of the other treatments. The later weaning treatments T18–T24 achieved higher survival rates (T18: 92.00 ± 2.27%; T21: 90.13 ± 2.06%; T24: 92.88 ± 5.20%), which were not significantly different from the Control.
._early_weaning_treatments_(t12-.jpeg)
Daily mortality patterns shown in Figure 2B indicate that fish in T12 began dying 2–3 days after weaning (15–16 DAH), with peak mortality at 23 DAH. In T15, mortality also began 2–3 days after weaning, with peak mortality observed at 18 DAH, but subsequently stabilized. In the later weaning treatments (T18–T24), mortality after weaning showed no sudden peaks, similar to the Control.
Growth
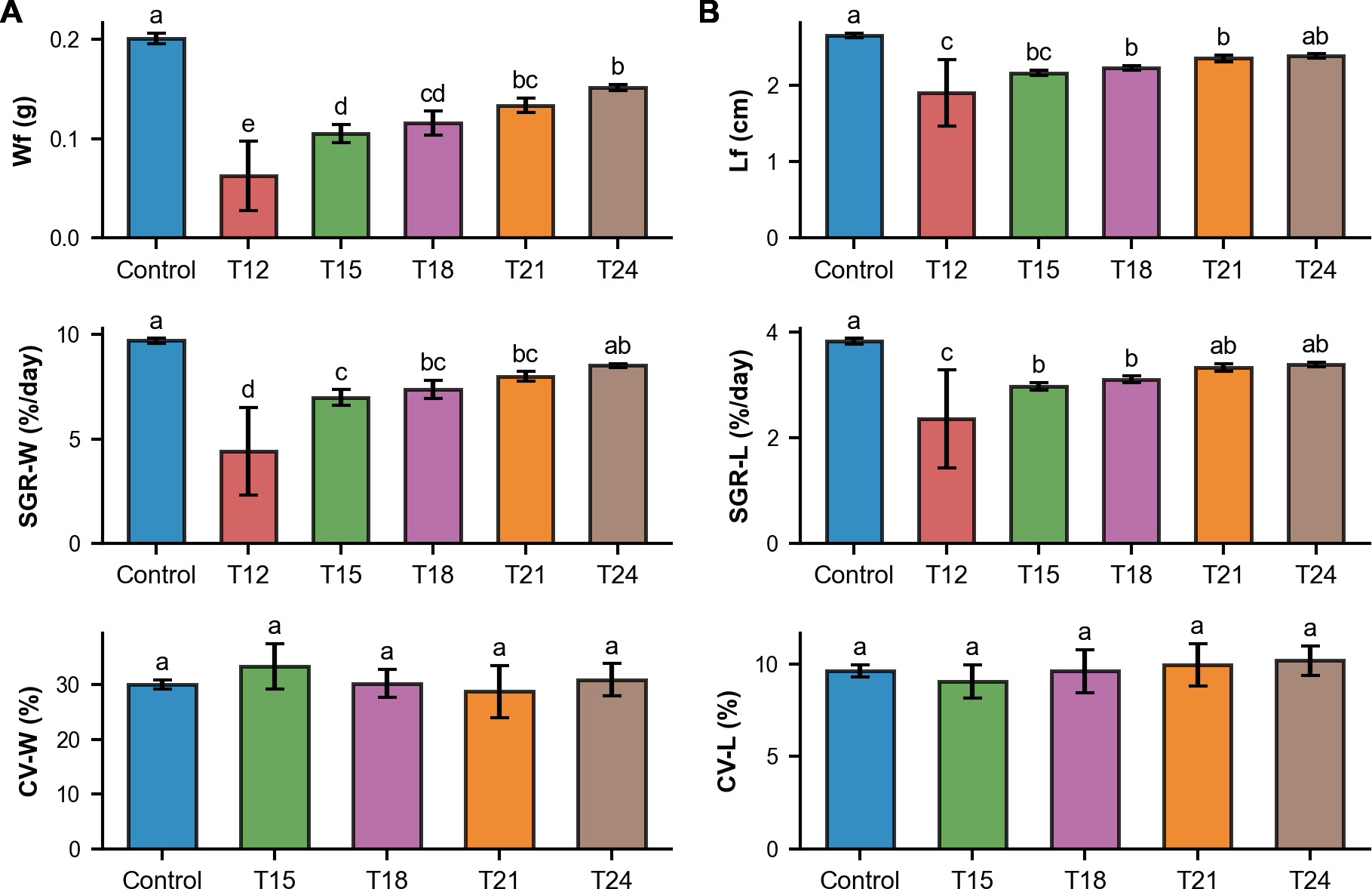
The growth of redtail botia increased with the later weaning treatments both in weight and length, see Figure 3. Overall, early weaning (T12–T15) resulted in reduced growth performance, whereas later weaning (T18–T24) produced outcomes comparable to the Control.
._early_weaning_treatments_(t12--t15)_had_the_lowest_growth_.png)
Final body weight (Wf) was significantly different among treatments (p<0.001). Early weaning treatments showed the lowest weights (T12: 0.062 ± 0.035 g; T15: 0.105 ± 0.009 g) while later weaning treatments produced higher weights (T18: 0.116 ± 0.012 g; T21: 0.134 ± 0.007 g); T24: 0.151 ± 0.003 g), closer to the Control (0.201 ± 0.006 g). The specific growth rate in weight (SGR-W) was significantly different among treatments (p<0.001). Early weaning treatments had the lowest (T12: 4.40 ± 2.10%/day; T15: 6.99 ± 0.37%/day) while later weaning treatments showed higher growth rates (T18: 7.38 ± 0.45%/day; T21: 7.99 ± 0.23%/day; T24: 8.52 ± 0.09%/day), closer to the Control (9.70 ± 0.11%/day). Coefficient of variation in weight (CV-W) was not significantly different among treatments (28.69%–33.28%). The CV-W value for T12 was considered unreliable due to the extremely low number of surviving fish.
Final body length (Lf) was also significantly different among treatments (p<0.001). Early weaning treatments showed the lowest lengths (T12: 1.90 ± 0.44 cm; T15: 2.16 ± 0.04 cm) while later weaning treatments produced higher weights (T18: 2.23 ± 0.03 cm; T21: 2.36 ± 0.04 cm); T24: 2.39 ± 0.03 cm), closer to the Control (2.66 ± 0.03 cm). The specific growth rate in length (SGR-L) was significantly different among treatments (p<0.01). Early weaning treatments had the lowest (T12: 2.36 ± 0.92%/day; T15: 2.98 ± 0.07%/day) while later weaning treatments showed higher growth rates (T18: 3.11 ± 0.06%/day; T21: 3.33 ± 0.08%/day; T24: 3.39 ± 0.05%/day), closer to the Control (3.83 ± 0.05%/day). Coefficient of variation in length (CV-L) was not significantly different among treatments (9.05%–10.17%). The CV-L value for T12 was considered unreliable due to the extremely low number of surviving fish.
Digestive enzyme activities
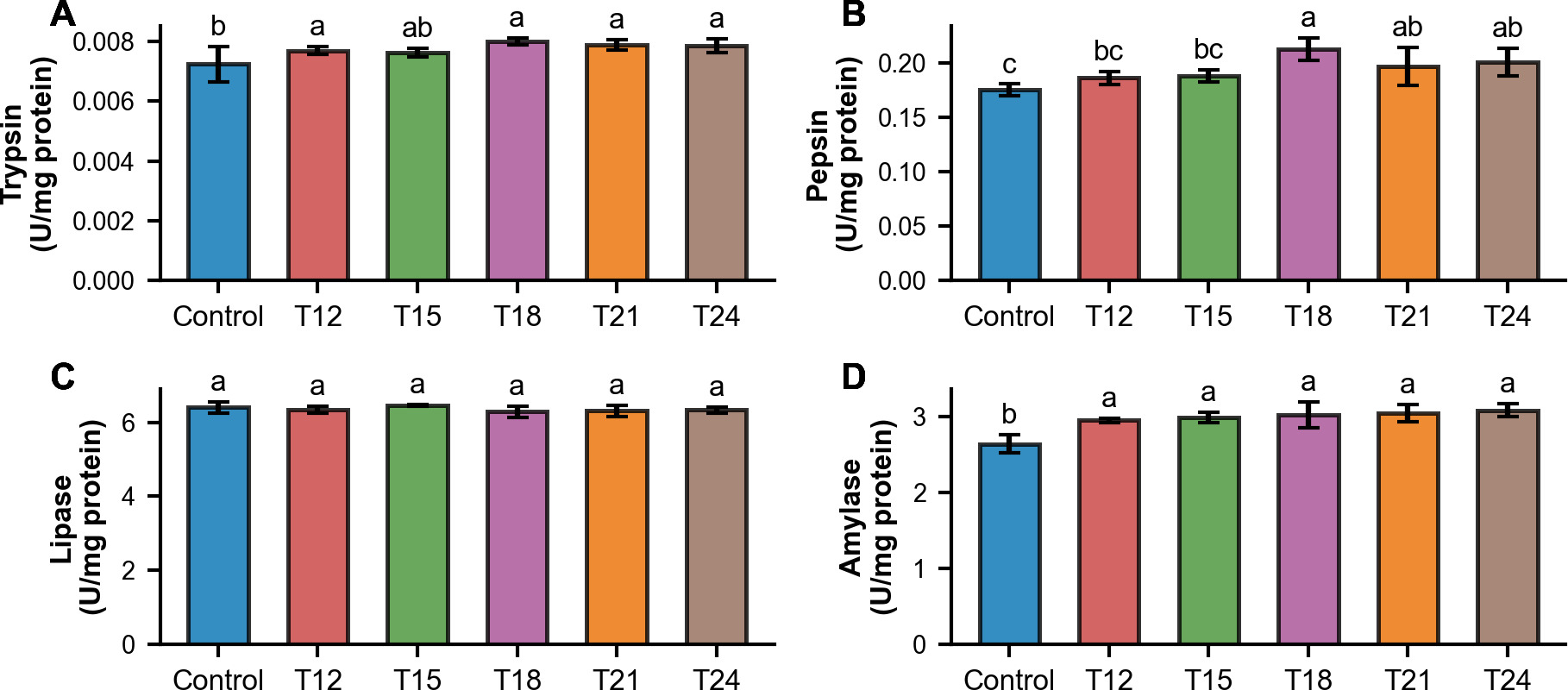
The digestive enzyme activities of redtail botia in the weaning treatments were higher than those in the Control, except for lipase (Figure 4). Trypsin activity was significantly affected by weaning time (p<0.05). The Control treatment showed the lowest value (0.0072 ± 0.0006 U/mg protein) and was significantly lower than the other treatments (T12: 0.0077 ± 0.0001; T18: 0.0080 ± 0.0001; T21: 0.0079 ± 0.0002; T24: 0.0079 ± 0.0002), except for T15 (0.0076 ± 0.0001). No significant differences were observed among the weaning treatments (T12–T24). Pepsin activity also differed significantly among treatments (p<0.01). The Control treatment showed the lowest value (0.175 ± 0.005 U/mg protein), which was significantly lower than the later weaning treatments (T18: 0.213 ± 0.010; T21: 0.197 ± 0.017; T24: 0.201 ± 0.013), but not the early weaning treatments (T12: 0.186 ± 0.006; T15: 0.188 ± 0.006).
._weaning_treatments_showed_higher_acti.png)
Lipase activity was not significantly influenced by weaning time (p = 0.346). Mean values ranged narrowly across treatments with no clear differences between the Control treatment (Control: 6.407 ± 0.155 U/mg protein) and the other weaning treatments (T12: 6.335 ± 0.092; T15: 6.461 ± 0.016; T18: 6.286 ± 0.149; T21: 6.315 ± 0.151; T24: 6.333 ± 0.084).
Amylase activity was significantly affected by weaning time (p<0.001). The Control treatment showed the lowest value (2.64 ± 0.12 U/mg protein) and was significantly lower than the other treatments (T12: 2.95 ± 0.03; T15: 2.99 ± 0.06; T18: 3.02 ± 0.18; T21: 3.04 ± 0.11; T24: 3.08 ± 0.08). No significant differences were observed among the weaning treatments (T12–T24).
Nutritional composition of fish
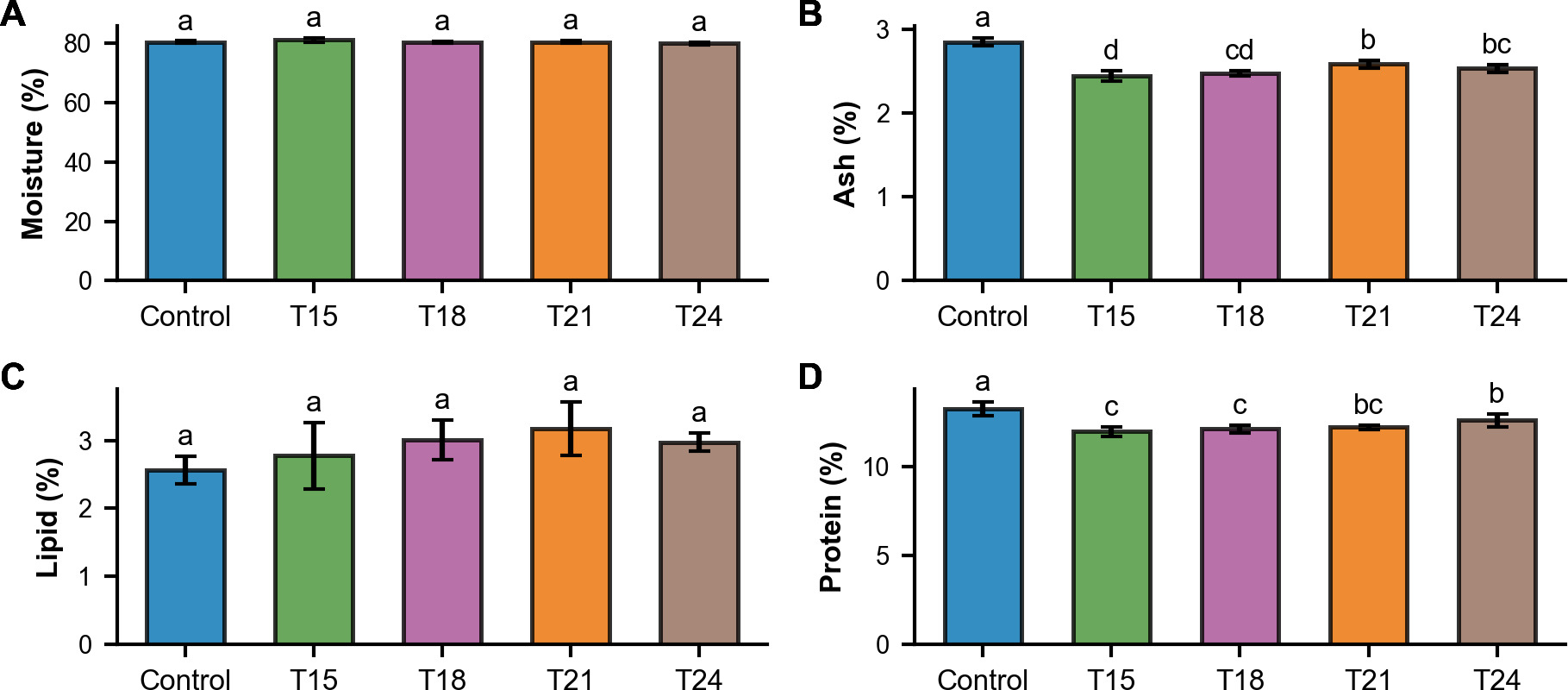
The nutritional composition of redtail botia varied among treatments (Figure 5). Due to the low number of surviving fish, nutritional analysis could not be performed for T12. Moisture content was not significantly influenced by weaning time (p = 0.052). Moisture values ranged from 80.0 ± 0.50% (T24) to 81.1 ± 0.65% (T15). Ash content was significantly affected by weaning time (p<0.01). The Control treatment had the highest ash value (2.85 ± 0.05%) and was significantly higher than those of the weaning treatments (T15: 2.45 ± 0.06%; T18: 2.48 ± 0.03%); T21: 2.59 ± 0.05%; T24: 2.54 ± 0.05%). Lipid content was higher in the weaning treatment with the highest value in T18 (3.01 ± 0.29%) compared to the Control (2.65 ± 0.13%). However, weaning time did not significantly influence the differences (p = 0.14). Protein content differed significantly among treatments (p<0.01). The Control treatment had the highest protein level (13.27 ± 0.39%) and was significantly higher than those of the weaning treatments. Earlier weaning treatments tended to have lower protein content (T15: 11.97 ± 0.28%; T18: 12.12 ± 0.23%) compared to the later weaning treatments (T21: 12.22 ± 0.12%; T24: 12.60 ± 0.37%).
._weaning_treatments_showed_lower_p.png)
Discussion
Survival and growth of redtail botia larvae were strongly influenced by weaning time, which is closely linked to the development of the digestive system and enzyme activities, consistent with findings in both freshwater and marine larvae.10,13,40,41 Early weaning (12–15 DAH) resulted in sharp declines in survival, with peak mortality occurring 2–3 days after the transition to formulated feed, and reduced growth in both weight and length. In contrast, later weaning (18–24 DAH) maintained high survival rates and growth comparable to feeding solely with live feed.
Larvae require a high protein hydrolysis efficiency to digest artificial diets effectively.42 Most fish larvae possess a rudimentary digestive system immediately after hatching, which limits their ability to absorb formulated feed.13 According to our previous study,4 the stomach of redtail botia is clearly differentiated by 7 DAH, and gastric glands are identifiable by 12 DAH. By 15 DAH, the digestive tract has completed its morphological development with the appearance of the pyloric sphincter, a constriction between the stomach and intestine. Regarding digestive enzymes, alkaline protease and amylase activities have been present since hatching, whereas pepsin activity has only been detectable at 12 DAH, but at a very low level. This may have resulted in lower feed intake caused by poor palatability and limited nutrient digestibility at the early larval stage. In addition, microdiets lack endogenous enzymes, forcing larvae to mobilize body reserves for energy production, which negatively affects growth.32 Therefore, weaning to formulated feed at 12–15 DAH seems to be too early for the digestive system of redtail botia. Such low survival rates under early weaning have also been reported in several freshwater catfish species, including Chinese long-snout catfish Leiocassis longirostris,30 butter catfish Ompok bimaculatus,28 and Clarias magur.31 Survival rates and growth of redtail botia under later weaning from 18 DAH improved and were comparable to feeding exclusively on Moina, indicating that by this stage, the digestive tract had sufficiently developed to support the transition to artificial diets. Delayed weaning, therefore, appears to support larval resilience and minimize mortality fluctuations.42–44
Weaning onto formulated feed had certain influences on the digestive enzyme activities of redtail botia larvae. In this study, enzyme activities increased in larvae weaned onto microdiets compared to those fed solely with Moina, except for Lipase. Enhanced digestive enzyme activity following weaning is often considered a compensatory adaptation to nutritional stress.45,46 Also, digestive enzyme activity in fish is age-dependent but can be modulated through dietary adjustments.18 Specifically, trypsin activity of redtail botia increased in larvae weaned onto microdiets compared to those fed exclusively on Moina, and no significant differences were observed among the different weaning times. A similar phenomenon in trypsin activity has been reported in Chinese longsnout catfish Leiocassis longirostris weaned onto microdiets compared to those fed solely on Artemia.30 For pepsin activity, early weaning (12–15 DAH) resulted in lower values compared to later weaning (18–24 DAH), but still higher than feeding solely with Moina. This may be explained by the fact that gastric glands appear only around day 12 DAH, when pepsin activity first becomes detectable.4 The gastric secretion converts pepsinogen into pepsin, initiating protein digestion.47 However, the presence of gastric glands does not necessarily indicate that the digestive tract is functionally mature, and larvae weaned at earlier stages likely lacked sufficient gastric digestive capacity to efficiently utilize formulated feed.48 Thus, weaning at this stage may impair pepsin activity by slowing stomach development.45 Amylase activity also followed a similar pattern to trypsin, increasing in weaning treatments compared to feeding solely with Moina, and no significant differences were observed among the different weaning times. Although amylase activity is considered genetically programmed and detectable at high levels during the yolk-sac stage, it can be stimulated by dietary carbohydrates, such as starch and glycogen, in microdiets.13,40,46,49 In contrast to the previous digestive enzymes, lipase activity was not significantly affected, which is consistent with findings in Chinese longsnout catfish30 and climbing perch.32 Collectively, the digestive enzyme activities of redtail botia across different weaning times indicate that, following an adaptation period to artificial diets, and once the digestive system is fully developed, larvae are capable of considerable flexibility in utilizing different feeding strategies.13,50,51
Weaning onto formulated feed had certain influences on the nutritional composition of redtail botia larvae. Previous studies showed that the nutritional composition of feed has a major influence on fish body composition, particularly protein and lipid levels, as well as ash (mineral content).32,52,53 We detected that the protein content of redtail botia was highest in the Moina treatment, significantly different from the microdiet weaning treatments. This can be explained by the higher protein content of Moina (71.7%) compared to microdiets (46.9%), as well as the higher efficiency of protein accumulation from live feed relative to formulated diets. We also observed the lowest lipid content in the Moina treatment. However, this was not statistically significant, likely due to the experiment’s limited statistical power. Similar findings have been reported across different species. In silver perch Bidyanus bidyanus, carcass protein content was positively correlated with dietary protein levels.52 In A. testudineus larvae, proximate composition varied significantly among treatments, with the lowest protein and highest lipid levels observed in groups fed formulated diets.32 Likewise, juvenile red drum fed commercial pellet diets exhibited higher body fat but lower protein and ash content compared to those fed live feed, suggesting that pelleted feeds may promote fat deposition while reducing protein and mineral accumulation, likely due to imbalances in nutrient composition between live feed and formulated diets.53
Our findings emphasize the importance of synchronizing weaning time with the development of digestive capacity to ensure both survival and growth, particularly survival, since growth can be compensated during later stages of rearing.54 Since live feed production is expensive, weaning fish as early as possible is a major economic challenge in aquaculture to reduce production costs.55 Our study suggests that the most appropriate weaning from live feed to formulated feed for redtail botia is 18 DAH, which is later than related catfish species, such as 10 DAH for Chinese long-snout catfish Leiocassis longirostris,30 7 DAH for butter catfish Ompok bimaculatus,28 15 DAH Clarias magur.31
Conclusion
This study investigated the effects of weaning redtail botia larvae from Moina to a formulated feed at different times. Weaning before 18 DAH was associated with a sharp decline in survival rates. These reduced survival rates are associated with the larvae’s immature digestive capacity and insufficient enzymatic activity. In contrast, by 18 DAH, the survival rate had improved, as the digestive tract had sufficiently developed to support the transition. Our findings suggest that the optimal weaning time for redtail botia to achieve high survival rates is around 18 DAH, at which survival and growth are comparable to those with continuous live feeding. Weaning onto formulated feed still influences digestive enzymes and nutritional composition. Further research is needed to improve diet formulation, enhance digestibility, and incorporate exogenous enzymes to support better growth outcomes during the transition from live feed to formulated feed.
Acknowledgment
The authors would like to thank the Bachelor of Aquaculture students (K46 and K47) at Can Tho University for their support throughout this study. This work was supported by the Ministry of Education and Training (MOET), Vietnam, under Grant Agreement No. B2024-TCT-12.
Author’s Contribution
Conceptualization: Tran T.T. Hien (Equal), Lam M. Lan (Equal). Data curation: Tran T.T. Hien (Equal), Lam M. Lan (Equal), Tran L.C. Tu (Equal), Nguyen H.Q. Thang (Equal), Dang T.H. Mai (Equal), Tran N.D. Khoa (Equal). Formal Analysis: Tran T.T. Hien (Equal), Dang T.H. Mai (Equal), Tran N.D. Khoa (Equal). Methodology: Tran T.T. Hien (Equal), Lam M. Lan (Equal). Writing – original draft: Tran T.T. Hien (Equal), Lam M. Lan (Equal). Writing – review & editing: Tran T.T. Hien (Equal), Lam M. Lan (Equal), Tran L.C. Tu (Equal), Nguyen H.Q. Thang (Equal), Dang T.H. Mai (Equal), Tran N.D. Khoa (Equal).
Ethical conduct approval
The animals used in this study were treated in accordance with institutional regulations under Decision No. 3965/QĐ-ĐHCT issued on October 15, 2021, titled “Can Tho University Regulation on Ethics in Animal Experimentation.”
Data availability
Data are available from the corresponding author upon reasonable request.
INFORMED CONSENT STATEMENT
All authors and institutions have confirmed this manuscript for publication.
Declaration of generative AI in scientific writing
During the preparation of this manuscript, the authors used Microsoft Copilot to enhance its readability and language quality. All content was thoroughly reviewed and edited by the authors, who take full responsibility for the content of the published article.