




Introduction
The whiteleg shrimp (Litopenaeus vannamei) is a dominant and economically important species in global aquaculture, accounting for a substantial proportion of global shrimp production.1 Its fast growth rate, high stocking-density culture, and broad environmental adaptability have made it a preferred species in many shrimp-farming regions, particularly in Asia and Latin America.2 However, shrimp aquaculture is facing increasing challenges, including disease outbreaks, environmental stress, and variable production performance. The intestinal microbiota also plays a crucial role in maintaining host physiological functions, including digestion, immune modulation, and protection against pathogenic microorganisms in aquatic animals.3,4 In shrimp, the gut microbial community is closely associated with growth performance, feed efficiency, and resistance to bacterial pathogens such as Vibrio spp. Environmental fluctuations, high stocking density, poor water quality, and excessive use of antibiotics can disrupt the balance of intestinal microbiota, leading to dysbiosis and increased disease risk.5,6 Therefore, the development of strategies that support a stable and beneficial gut microbial community has become a critical aspect of sustainable shrimp aquaculture. Among the various strategies proposed to enhance gut health and disease resistance in shrimp, functional feed additives, including probiotics, prebiotics, and synbiotics, have attracted increasing attention.
Prebiotics are defined as non-digestible feed ingredients that selectively stimulate the growth and activity of beneficial microorganisms in the gastrointestinal tract, thereby conferring health benefits to the host.7 In recent years, natural prebiotic extracts derived from plants, vegetables, algae, and agricultural by-products have emerged as promising candidates for aquaculture applications due to their safety, biodegradability, cost-effectiveness and additional bioactive properties. These extracts contain functional compounds such as oligosaccharides, polysaccharides, phenolic compounds, and dietary fibers that can serve as substrates for beneficial intestinal bacteria.8,9 For example, banana extracts contain high levels of polysaccharides and other bioactive compounds that may promote the growth and activity of Lactobacillus spp.10,11 Therefore, the development of a combination of prebiotics derived from natural extracts and probiotic bacteria, known as synbiotics, represents a promising and sustainable approach for improving growth, survival, digestive enzyme activity and disease resistance in aquacultured species. In shrimp, dietary supplementation with synbiotic extracts derived from natural products has been reported to improve growth performance, immune responses, antioxidant capacity, and resistance to bacterial infections. For example, Phan et al.12 reported that dietary supplementation with 1% noni leaf extract enhanced immune functions in whiteleg shrimp (L. vannamei). Situmorang et al.13 demonstrated that a synbiotic containing algae improved growth performance and survival of shrimp after challenge with Vibrio harveyi. However, the mechanisms underlying these beneficial effects are not yet fully understood, particularly with regard to their influence on the intestinal bacterial community, diversity, and functional dynamics.
Oral administration via feed supplementation is widely recognized as one of the most practical and effective strategies for delivering functional additives to shrimp.14,15 This approach enables direct interaction between dietary components and the gastrointestinal tract, where modulation of the microbial community primarily occurs. Accordingly, the present study investigated the effects of dietary supplementation with natural prebiotic extracts on the intestinal bacterial community of L. vannamei. By assessing microbial diversity and community composition, the objective of study is to clarify the role of natural prebiotics in modulating gut microbiota and to generate scientific evidence supporting their application as sustainable feed additives in shrimp aquaculture. Specifically, the study was designed with two complementary experiments: an in vitro experiment (Experiment 1) to evaluate the functional effects of the tested substrates, and an in vivo experiment (Experiment 2) to investigate microbial diversity and community composition in L. vannamei. The results are expected to enhance current understanding of diet–microbiota interactions in L. vannamei and to promote environmentally friendly approaches for improving shrimp health and production performance.
Materials and Methods
Experiment 1. Evaluation of prebiotic scores of natural extracts
Preparation of natural extracts: Prebiotic extract from banana was prepared following the methods of Lingyun et al.16 and Boonmee and Rengpipat.17 Ripe bananas were washed, peeled, sliced, and homogenized in hot water, then centrifuged. The supernatant was filtered, residues were re-extracted, and filtrates combined before concentration by rotary evaporation. The extract was freeze-dried and kept at −20°C until use.
Prebiotic extracts from peel of watermelon, muskmelon and papaya were prepared following the method described by Hao et al.18 For sweet potato, the extraction method described by Marlida et al.19 was used. All extracts were freeze-dried, ground into fine powder, and stored at −20 °C until further analysis.
Determination of prebiotic index: Prebiotic index was evaluated following the method of Mazzola et al.,20 with some modifications. The probiotic strain Lactobacillus plantarum LAB8.1CM was isolated from shrimp’s intestine and cultured in MRS medium supplemented with either 2% glucose or 2% prebiotic substrates (commercial FOS: RM9582, Himedia, India; and extracted products) as carbon sources. The optical density (OD) was measured at 0 and 24 h, and the prebiotic value was calculated using the formula of Mazzola et al.20 Each treatment was conducted in triplicate. The Lactobacillus plantarum LAB8.1CM exhibiting the highest prebiotic index when cultured with the natural extracts was selected for subsequent experiments. The strains of Vibrio parahaemolyticus and Vibrio harveyi used in this study were obtained from the Probiotic Laboratory, as described by Giang et al.21
Prebiotic index =
where ODi and ODt are the optical density values measured at 600 nm at 0 h and after 24 h of incubation, respectively; Lac.-Pre. and Lac.-Glu represent the culture media for Lactobacillus supplemented with the natural extracts and glucose, respectively; and Vib.-Pre. and Vib.-Glu. represent the culture media for Vibrio spp. supplemented with the extracted products and glucose, respectively.
Experiment 2. Effects of banana extract on alterations in the intestinal microbiota of whiteleg shrimp
Diet preparation: The probiotic strain Lactobacillus plantarum LAB8.1CM, isolated from whiteleg shrimp, was reactivated in MRS medium and mixed with skim milk powder to obtain a concentration of 10⁸ CFU/g, and stored at 4°C until use.
A commercial shrimp diet was supplemented with L. plantarum (10⁸ CFU/kg) and banana extract (BE) by spraying their saline suspensions onto pellets, followed by coating with 5% CMC and drying at 37°C. The LAB concentration in feeds was determined again by spread plate method.22 Prepared diets were stored at −20°C, with new batches produced weekly to maintain probiotic viability.
Experimental design: Post-larval shrimp were purchased from a commercial hatchery in Can Tho city, Vietnam. They were cultured at 4 m3 until use for the experiment. A total of 100 healthy shrimp (4.4 ± 0.3 g) were randomly distributed into six 0.5 m³ tanks (two treatments with triplicates). The experiment consisted of two dietary treatments: (1) a control diet (Ctrl) and a synbiotic diet (BE 0.2% + LAB8.1CM at 10⁸ CFU/kg; denoted as Syn thereafter). Shrimp were fed the experimental diets for 30 days at a feeding rate of 3–6% of body weight per day. Feed was divided three times per day. Uneaten feed was removed after 2 hours of feeding and tanks were siphoned every two days to remove accumulated waste.
Collection of intestinal samples and DNA extraction: After the 30-day feeding trial, six shrimp were randomly selected from each replicate (18 shrimp per treatment), and starved for 24 h prior to sampling. Shrimp were anesthetized in ice water, and intestinal samples were collected for DNA extraction using the DNeasy PowerSoil Pro Kit (Qiagen, USA) according to the manufacturer’s instructions. DNA concentration was measured using Qubit fluorometry, and purity was assessed by spectrophotometry (OD260/OD280). Samples with DNA concentration ≥ 0.20 ng/µL, total DNA ≥ 15.00 ng, and OD260/OD280 ≥ 1.50 were used for subsequent analyses.
16S metagenomic sequencing and data processing: The V3–V4 region of the 16S rRNA gene was amplified using universal bacterial primers 341F (5′-CCTACGGGNGGCWGCAG-3′) and 805R (5′-GACTACHVGGGTATCTAATCC-3′), followed by tag pyrosequencing.23 Non-chimeric sequences were analyzed using QIIME (v1.8.0), with closed-reference OTU picking against the Greengenes database (gg_13_8) at 97% identity.24 Pairwise distances were calculated with a cutoff of 0.03, and sequences were clustered into OTUs using the average neighbor algorithm. Representative OTUs were then taxonomically assigned. The identified taxa were further validated using the Global Catalogue of Microorganisms.25
The singleton sequences were removed.26 At the taxonomic levels from phylum to species and the result of clustering was visualized on principal components analysis (PCA) plot. In addition to PCA, heatmap representation of species-level bacterial composition, the relative abundance of genera for samples were also performed using a web tool for visualizing clustering of multivariate data (http://biit.cs.ut.ee/clustvis/).27
Indices of microbial community diversity, including species, number of species, Margalef’s species richness (d), Similarity, PCA eigen-vector plots were performed using Plymouth Routines in Multivariate Ecological Research (PRIMER) ver. 6.1.5.28
Statistical analysis
Results in experiment 1 are expressed as mean ± standard error (SE). Statistical analyses were performed using IBM SPSS Statistics 22.0. Treatment means were compared by one-way ANOVA followed by Tukey’s test (p < 0.05). For experiment 2, diversity indices were analyzed using an independent samples t-test (p < 0.05), based on replicate values (n = 3).
Results
Experiment 1. Determination of prebiotic scores of natural extracts: Pathogenic bacteria were used as reference strains, and prebiotic scores were calculated as the mean values obtained for Lactiplantibacillus plantarum LAB8.1CM. In Figures 1, FOS exhibited the highest prebiotic index, indicating the strongest prebiotic potential, followed by banana and sweet potato, while papaya and watermelon extracts showed significantly lower values.
_and_*v._harveyi*_(b.svg)
Experiment 2. Effects of banana extract on alterations in the intestinal microbiota of whiteleg shrimp
Water quality parameters remained stable in both the control (Ctrl) and synbiotic (Syn) treatments throughout the experimental period. No significant differences were observed in any measured parameters between the control and synbiotic treatments.
General intestinal microbiome characteristics: In general, 286 OTUs were obtained. After removing singletons, they were assigned to 8 phyla, 11 classes, 29 orders, 37 families and 63 genera with taxonomical names (Figure. 2).

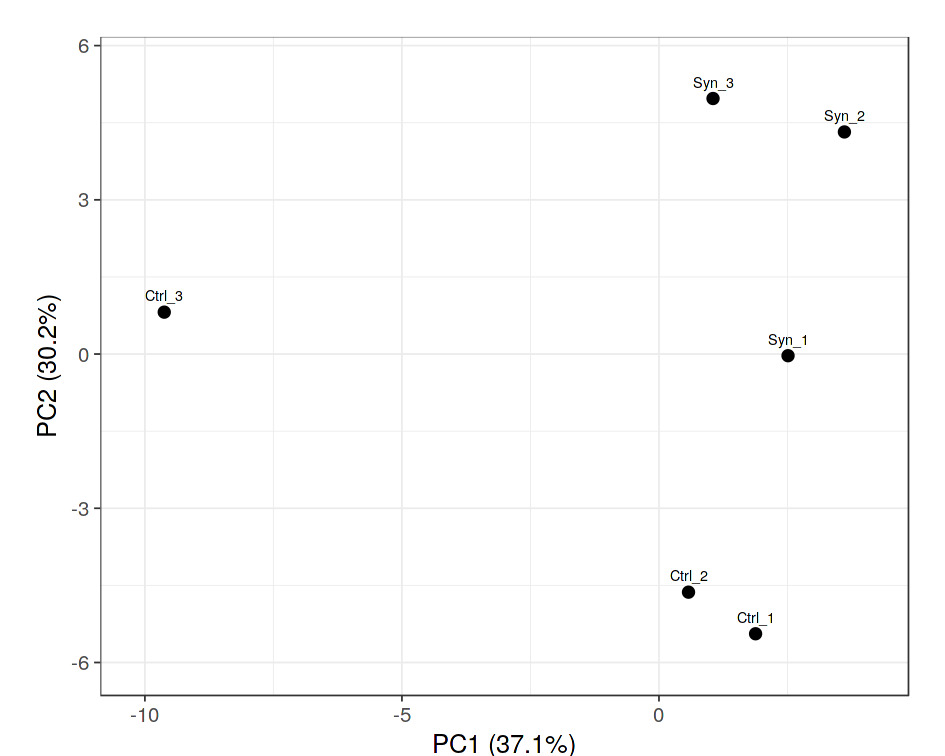
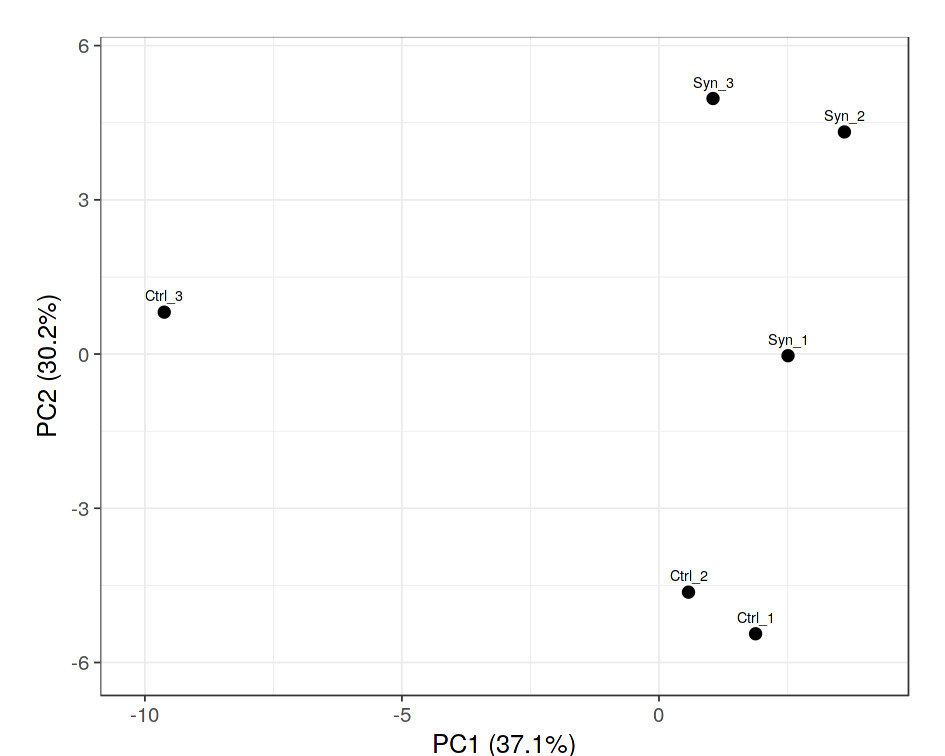
Synbiotic changed shrimp microbiome structure and diversity: The PCA analysis of the microbiota revealed the shrimp clustered into two distinct groups depending on diet. The contributions of PC1 and PC2 were 37.1% and 30.2%, respectively; explained together 67.3% of the variation in the dateset (Figure 3). Here, the bacteria explained the highest degree of variation belonged almost to phyla Proteobacteria, Bacteroidota, Actinobacteriota and Firmicutes. Relative abundance of Proteobacteria phylum in the intestine of L. vannamei that being fed the synbiotic and control diets were 90.2% and 76.8%, respectively. Interestingly, relative abundance of Firmicutes phylum were 5.8% and 1.5% synbiotic and control shrimp, respectively (Figure 4).

In general, at the genus level, the most abundant genera in the intestine of shrimp were Vibrio and Paracoccus. The relative abundance of the predominant genera differed between the control and synbiotic treatments. Six predominant genera, including Vibrio, Paracoccus, Shewanella, Ruegeria, Leisingera, and Enterobacter were detected in control shrimp, whereas 10 predominant genera, including Vibrio, Paracoccus, Pseudoalteromonas, Ruegeria, Lactobacillus, Algoriphagus, Bacillus, Sulfitobacter, Leisingera, and Enterobacter were found in SYN-fed shrimp (Figure 5). Hierarchy cluster heatmap analysis intuitively confirmed high or low genus in each intestinal sample (Figure 6).
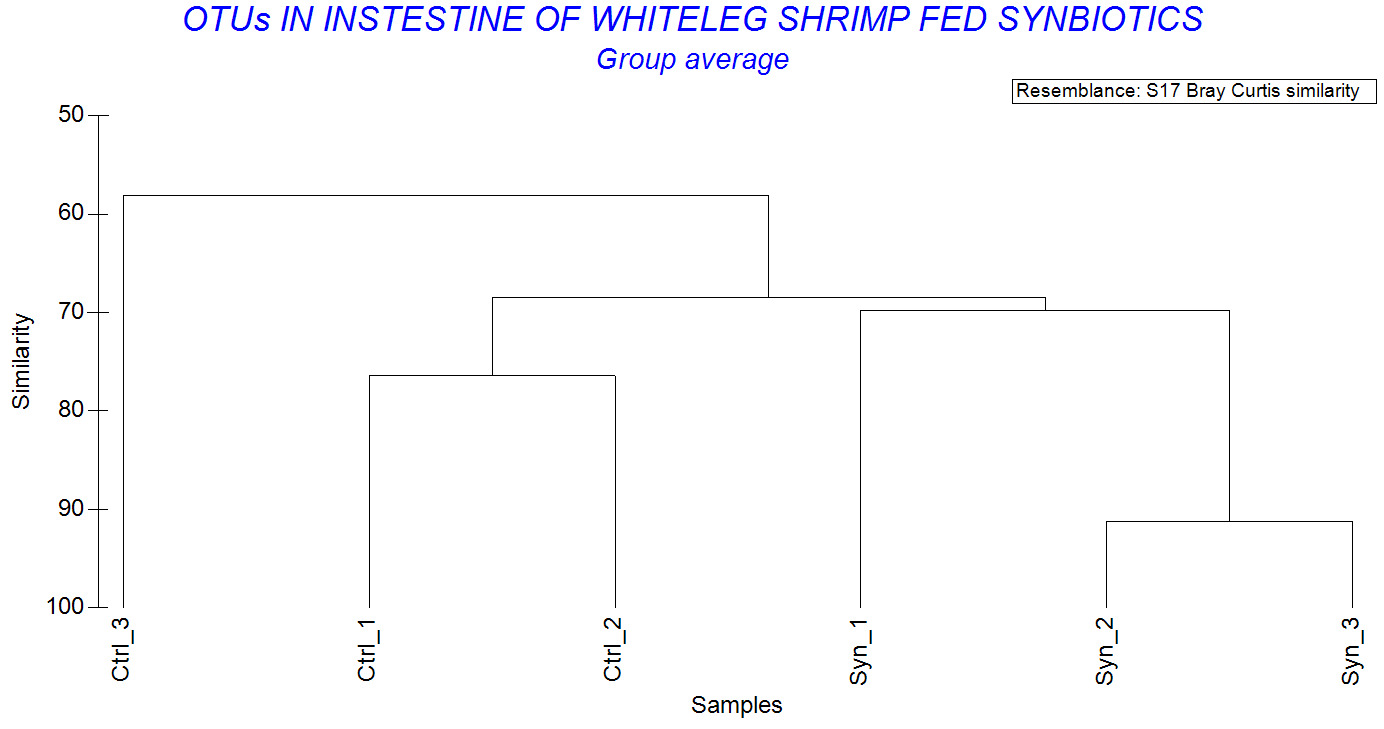
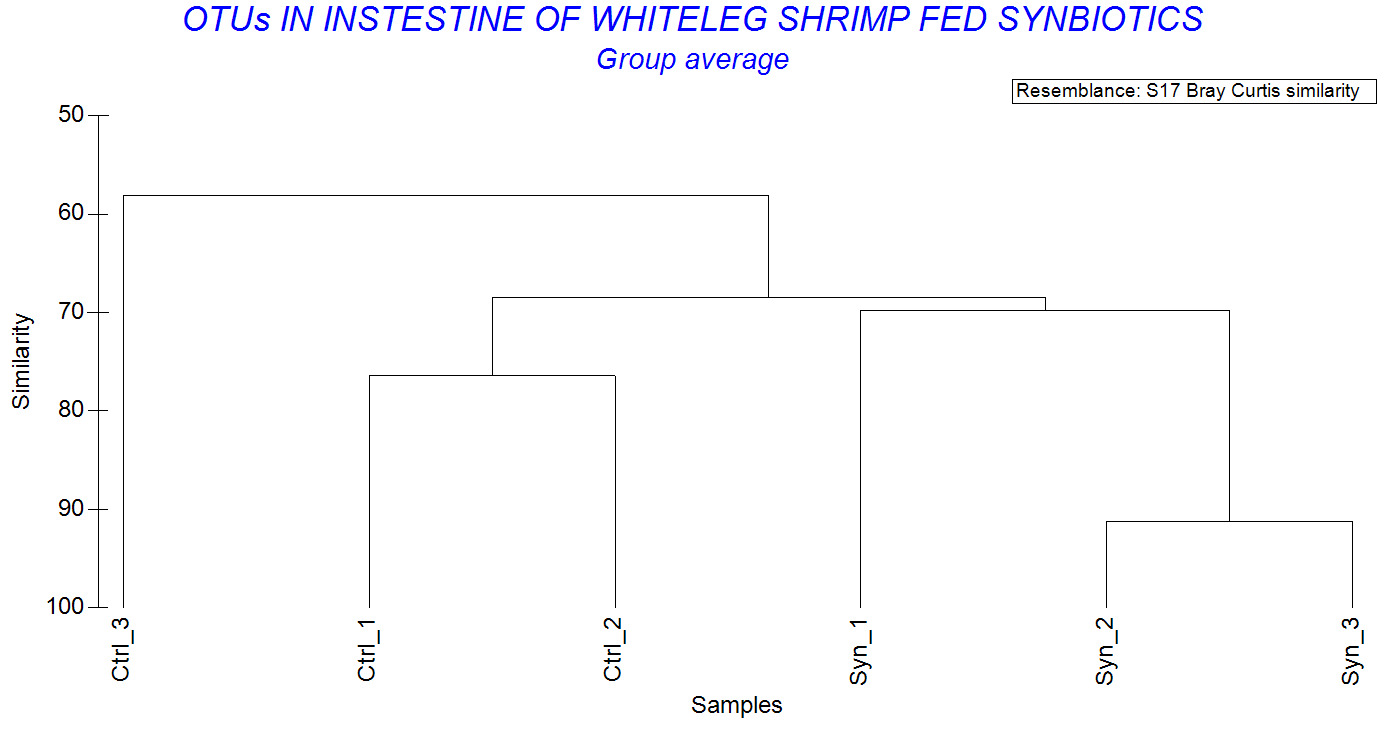
The Biodiversity similarity analysis showed that Syn_2 and Syn_3 had a Bray Curtis similarity index of 91.2%, while Syn_1 and Syn_2 were 70.7%. In contrast, the similarity indices of control shrimp were low, especially Ctrl_2 and Ctrl_3 was only 56.7% (Figure 7). Overall, within-group similarity tended to be higher in the synbiotic treatment compared to the control group, indicating a more consistent microbial community structure.


Biodiversity Indices
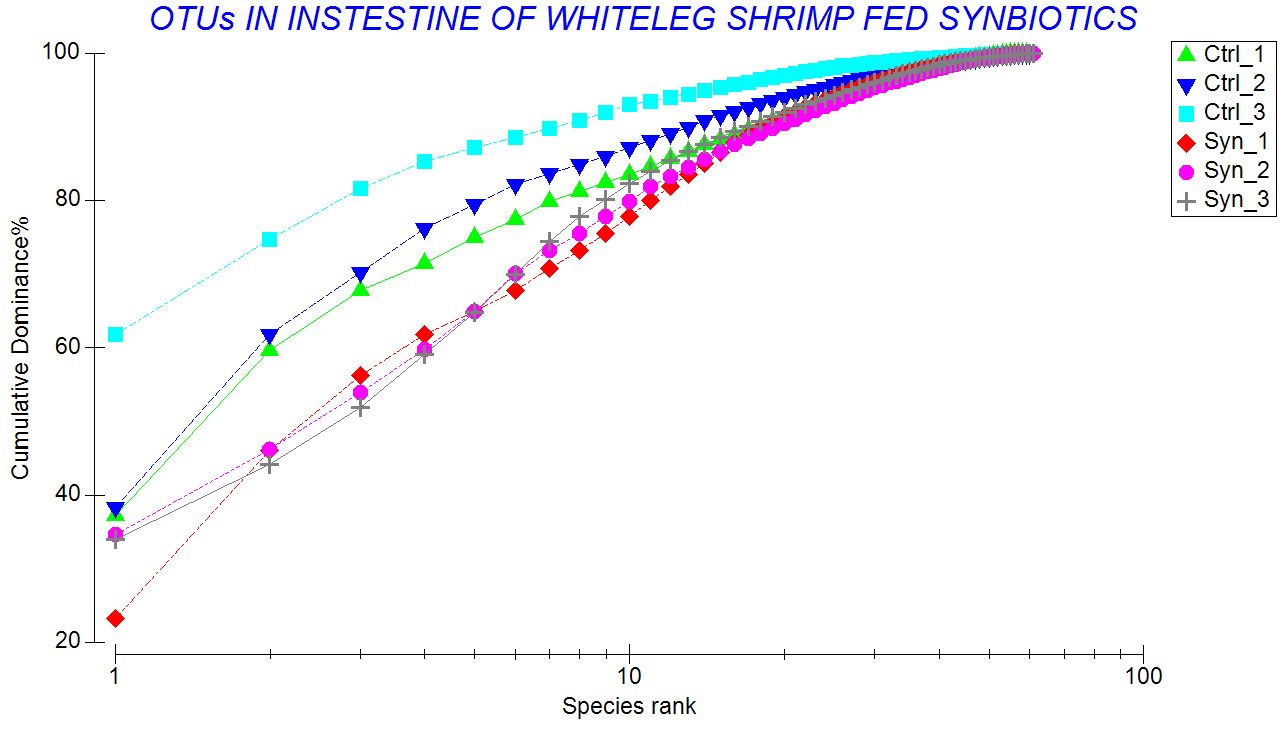
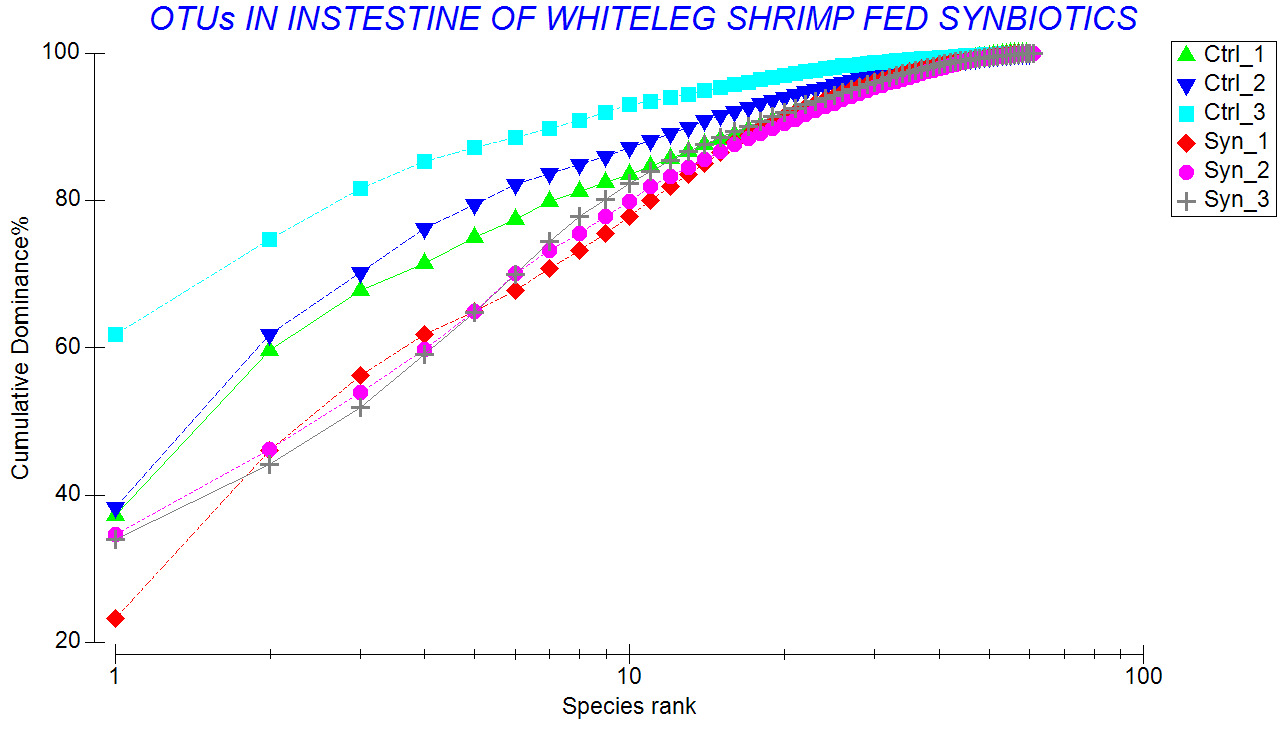
The results showed that shrimp fed the SYN diet had higher species richness, and Shannon-Weaver index than those of the control shrimp (Table 2). Lower Pielou’s evenness (J’) indicates that several species dominated in the control shrimp. This was consistent with those shown in the k-dominance curves (Figure 8) and percentage of relative abundance (Figure 5).
Discussion
The present study demonstrates that dietary supplementation with banana-derived prebiotic extract, especially when combined with L. plantarum LAB8.1CM, significantly enhances the beneficial modulation of the intestinal microbiota in L. vannamei. By integrating in vitro prebiotic screening and in vivo feeding trials with high-throughput 16S rRNA sequencing, this study provides deeper insights into diet–microbiota interactions in shrimp aquaculture and supports the development of synbiotic-based functional feeds as sustainable alternatives to antibiotics. The first experiment demonstrated that banana extract exhibited one of the highest prebiotic index values among the tested natural extracts, ranking second only to fructooligosaccharides (FOS), a well-established commercial prebiotic. The prebiotic index reflects the selective stimulation of beneficial bacteria relative to pathogenic strains, particularly Vibrio spp., which are major opportunistic pathogens in shrimp aquaculture. The strong performance of banana extract may be attributed to its high content of non-digestible carbohydrates, including resistant starch, inulin-like fructans, and other oligosaccharides, as well as phenolic compounds that can influence microbial metabolism.10,11 The selective stimulation of L. plantarum LAB8.1CM suggests that banana-derived substrates act as fermentable carbon sources for lactic acid bacteria, whose metabolic activity produces organic acids, bacteriocins, and other antimicrobial compounds that inhibit pathogenic bacteria. Such selective enhancement is particularly important in aquaculture systems, where dysbiosis and pathogen overgrowth are commonly induced by environmental stressors, including high stocking density, fluctuations in water quality, and nutritional imbalances.5,6 Therefore, the in vitro findings supported the selection of banana extract for subsequent in vivo evaluation as a component of the synbiotic formulation.
The feeding trial clearly demonstrated that dietary supplementation with banana extract combined with L. plantarum significantly influenced the structure of the intestinal microbial community. Principal component analysis (PCA) revealed distinct clustering between synbiotic-fed shrimp and the control shrimp, with PC1 and PC2 collectively accounting for 67.3% of the total variance. This clear separation indicates that dietary intervention was a primary factor shaping gut microbiota composition. At the phylum level, Proteobacteria remained the dominant phylum in both treatments, accounting for 90.2% in control shrimp and 76.8 % in synbiotic-fed shrimp. The predominance of Proteobacteria in shrimp intestines has been consistently reported and is considered characteristic of marine invertebrate microbiomes.5 However, an important finding of this study was the increased relative abundance of Firmicutes in synbiotic-fed shrimp (5.8%) compared to control shrimp (1.5%). Since many probiotic genera, including Lactobacillus and Bacillus, belong to the phylum Firmicutes, this shift suggests successful stimulation and/or colonization of beneficial bacteria. The increased abundance of Firmicutes may also indicate an enhanced capacity for carbohydrate fermentation in the intestine. Members of this phylum are well recognized for their ability to metabolize complex polysaccharides into short-chain fatty acids (SCFAs), which contribute to improved epithelial integrity, modulation of host immune responses, and enhanced nutrient absorption.29–31
Regarding genus level, a significant difference was observed between treatments. While Vibrio and Paracoccus were predominant in both groups, synbiotic-fed shrimp exhibited a broader spectrum of predominant genera, including Lactobacillus, Bacillus, Pseudoalteromonas, Algoriphagus, and Sulfitobacter. The detection of Lactobacillus in synbiotic shrimp suggested that supplemented L. plantarum strain was capable of transient persistence within the gastrointestinal tract. A similar finding has been reported previously.26 In that study, synbiotic supplementation with Lactobacillus plantarum combined with galactooligosaccharides significantly modulated the gut microbial community of shrimp, increasing the abundance of beneficial taxa and enhancing disease resistance.26 Moreover, shrimp fed the synbiotic diet exhibited higher Margalef’s species richness (d), Shannon–Wiener diversity (H’), and Pielou’s evenness (J’) compared to control shrimp. Increased diversity is generally considered a hallmark of a stable and resilient microbial ecosystem. Greater species richness may enhance functional redundancy, allowing the microbial community to maintain functional stability under environmental stress.32,33 In contrast, the control group exhibited lower community evenness and stronger dominance by a limited number of taxa, as illustrated by the k-dominance curves. Such dominance patterns may increase susceptibility to environmental perturbations and opportunistic pathogen invasion. Furthermore, the higher Bray–Curtis similarity observed among synbiotic replicates (up to 91.2%) suggests reduced inter-individual variability and a more consistent microbial community structure. This enhanced stability and uniformity are particularly important in intensive aquaculture systems, where fluctuating environmental conditions can rapidly disrupt microbial balance and compromise host health. These findings align with the concept that functional feed additives can shape gut microbial ecology in a predictable and beneficial manner.4 By providing selective substrates and beneficial microorganisms simultaneously, synbiotics may exert synergistic effects that exceed those of probiotics or prebiotics alone. Furthermore, modulation of the gut microbiota may have important implications for host immune function. Lactic acid bacteria are widely recognized for their ability to stimulate innate immune responses, enhance hemocyte activity, and improve resistance to bacterial pathogens.7,15 A well-balanced microbial community can also reinforce gut barrier integrity, thereby limiting pathogen translocation and reducing the risk of systemic infection.30,31 Although immune parameters were not comprehensively assessed in the present study, the microbial profile observed in synbiotic-fed shrimp indicates a more favorable intestinal environment that may contribute to enhanced disease resistance.
In summary, dietary supplementation with banana extract in combination with L. plantarum LAB8.1CM effectively reshaped the intestinal microbiota of L. vannamei. The synbiotic approach enhanced microbial diversity, increased the abundance of beneficial taxa such as Firmicutes and Lactobacillus, promoted community stability, and reduced dominance patterns associated with dysbiosis. These microbiological changes likely contributed to improved digestive performance and overall shrimp health. This study highlights the potential of natural prebiotic extracts as sustainable feed additives and provides valuable insights into diet–microbiota interactions in shrimp aquaculture. Future research should further explore functional metabolic outputs, immune responses, and pathogen challenge models to fully elucidate the mechanisms underlying synbiotic-mediated health benefits.
Acknowledgments
This work was funded by the Vietnamese Ministry of Education and Training (Project No. B2024-TCT-03). We appreciate Misses Diem My Ho, and Pham Yen Ngoc Nguyen for their assistances with the experiment.
Authors’ Contribution
Conceptualization: Giang T. Huynh (Equal). Methodology: Tu T.C. Phan (Equal), Giang T. Huynh (Equal), Phu Q. Truong (Equal). Formal Analysis: Giang T. Huynh (Equal), Hai H. Vu (Equal). Writing – original draft: Giang T. Huynh (Lead), Tu T.C. Phan (Equal). Writing – review & editing: Tu T.C. Phan (Equal), Giang T. Huynh (Equal). Funding acquisition: Giang T. Huynh (Lead).
Ethical Conduct Approval – IACUC
All animal procedures complied with Vietnam’s Law on Animal Health (2015; No. 79/2015/QH13).