1. INTRODUCTION
Chinese mitten crab (Eriocheir sinensis), an endemic economic freshwater crustacean species native to China, is widely recognized for its palatable flesh and prominent nutritional value. Facilitated by the continuous advancement of aquaculture technologies and the escalating market demand, the production of E. sinensis has exhibited a consistent growth trajectory over the recent decade. According to the China Fishery Statistical Yearbook (2023),1 the annual yield of this species increased from 593,000 tons in 2010 to 889,000 tons in 2023, accounting for a cumulative growth rate of 49.9%. However, as the scale and intensity of crab farming continue to expand, issues such as reduced stress resistance, slowed growth, and frequent disease outbreaks, including saprolegniosis, black gill disease, and rotten limb disease, have become increasingly severe due to high-density cultivation and deteriorating water quality.2 These problems significantly compromise farming profitability. To prevent and control these diseases, antibiotics and chemicals are often used in aquaculture production. However, the long-term application of these agents has led to enhanced drug resistance in pathogens, weakened immunity in crabs, and potential risks of drug residues.3 Consequently, identifying eco-friendly alternatives for disease prevention and immune enhancement has become a critical issue for achieving sustainable, healthy aquaculture in the Chinese mitten crab industry. In recent years, a variety of immunostimulants and immunomodulators have been shown to effectively prevent diseases, including β-glucan,4 peptidoglycan,5 chitin,6 vitamins,7 and various animal and plant products.8 These substances exert their effects by enhancing non-specific cellular defense mechanisms and humoral immune responses. Chitin is one of the most widespread complex polysaccharides in nature after cellulose. It is composed of N-acetylglucosamine (GlcNAc) linked by β-1,4-glycosidic bonds and is a structural homologue of cellulose.9 Chitin is widely distributed in the cell walls of fungi,10 the exoskeletons of insects.11 In addition, chitin is also an important component of the exoskeletons and peritrophic matrices of aquatic crustaceans.12
Current studies have shown that chitin plays an important role in enhancing various physiological functions of aquatic animals, including improvements in growth performance,13 enhanced immunity, and increased resistance against bacterial infections.6 Research by Hu et al.14 on largemouth bass (Micropterus salmoides) revealed that dietary supplementation with both black soldier fly larvae chitin and shrimp chitin significantly enhanced non-specific immune function and reduced the abundance of potential pathogenic bacteria in the intestine. Cheng et al.6 also identified chitin as a feasible immunostimulant that markedly upregulates immune enzyme activities and the expression of immune-related genes in Pacific white shrimp (Litopenaeus vannamei). Harikrishnan et al.15 demonstrated that dietary chitin supplementation could effectively enhance immune response activity of brown-marbled grouper (Epinephelus bruneus) and improve its resistance to infections by Philasterides dicentrarchi and Vibrio alginolyticus. Similar findings have been reported in gilthead seabream (Sparus aurata L.)16 and Micropterus salmoides.14
As a typical crustacean, E. sinensis undergoes multiple molting events during its growth and frequently consumes its own exuviae after molting. This behavior implies that chitin from the exoskeleton enters the digestive tract, where endogenously secreted chitinases can degrade it and subsequently reuse it. Compared with most fish species, which either lack endogenous chitinase secretion or exhibit low chitinase activity, E. sinensis utilizes chitin more directly and efficiently. However, no studies have reported the application of chitin in the diets of E. sinensis, and the appropriate supplementation level and functional effects of chitin remain unclear. To this end, different chitin concentrations were added to the control diet to evaluate their effects on the growth performance and immune function of E. sinensis. This study aims to provide a certain theoretical basis and practical reference for the application of dietary chitin on E. sinensis.
2. MATERIALS AND METHODS
2.1. EXPERIMENTAL DIETS AND DESIGN
The diet without chitin was the control diet (C1), which was designed to contain 200 g/kg fish meal, and then, 0.7% (C2), 1.4% (C3), 2.1% (C4), and 2.8% (C5) chitin were supplemented in the control diet to form five isoproteic and isolipidic diets, respectively. All ingredients were thoroughly blended according to the formulation in Table 1. This mixture was homogenized with fish oil and distilled water, then extruded into sinking pellets (pelleting temperature of 85 ± 5°C) using a single-screw extruder (SLP-22D, Dantu district Xin Fenghao Construction Machinery Factory). The pellets were post-cooked in an oven at 95°C for 20 min, then air-dried, and stored at room temperature until use.
Fish meal, DYM, meat and bone meal, soybean meal, peanut meal, cottonseed meal, and brewers dried yeast were the main protein sources in the diet. Fish oil and soyabean lecithin were added to meet fat needs, and flour and corn were added to meet the carbohydrate needs of E. sinensis. The Fish meal and fish oil were purchased from a provender mill in Taizhou, Jiangsu, China. The chitin used in the experiment was purchased from Shanghai Macklin Biochemical Technology Co., Ltd., derived from crab shells with an effective content of 99.9%.
2.2. EXPERIMENTAL CRAB AND FEEDING MANAGEMENT
Juvenile E. sinensis were obtained from Jiangsu Haorun Biological Industry Group Co., Ltd. (Jiangsu, China). All crabs were acclimated in a canvas pool (3 m×5 m×1.2 m) with crab nests as shelters and circulating water. During acclimation, the water temperature, dissolved oxygen, pH, ammonia nitrogen, and nitrite were kept at 23-28 ℃, ≥5.6 mg L-1, 7.0-8.5, ≤0.2 mg L-1, and ≤0.05 mg L-1, respectively. The crabs were fed the C1 diet at 07:00 and 17:00 each day for two weeks. Following an acclimation period, a total of 400 juvenile crabs (1.74 ± 0.10 g) in the intermolt period with vigorous appendages were selected. These crabs were randomly assigned to five groups, each replicated in quadruplicate. The feeding trial took place in the PVC tanks (1.2 m × 1.2 m × 0.75 m), with a crab nest as a shelter and circulating water.
During the 56-day feeding period, the crabs were fed twice daily at 07:00 and 17:00. The daily feed intake was approximately 1-3% of body weight. The feed intake was appropriately adjusted based on the water temperature, weather, and feeding behavior of the crabs to ensure the diets could be eaten up in 4 h after feeding. During the cultivation period, the sediment and faeces were removed by siphoning every day, and settled river water was supplemented. The water temperature, dissolved oxygen, pH, ammonia nitrogen, and nitrite were kept at 23-28 ℃, ≥5.6 mgL-1, 7.0-8.5, ≤0.2 mgL-1, and ≤0.05 mgL-1, respectively.
2.3. MEASUREMENT INDICATORS AND METHODS
At the end of the experiment, sample collection and the measurement of all indicators, including growth performance, proximate composition of feed and crab, serum biochemical indices, digestive enzyme activities, intestinal histology and microbial study were conducted in accordance with the experimental methods described in our previously published articles.17
After 24 h of fasting at the end of the experiment, the crabs in each tank were counted and weighed to determine survival, weight gain, and feed conversion ratio. Three crabs in each tank were randomly collected for the entire crab body composition analysis. After anesthetizing the juvenile crabs in ice water, four crabs were randomly selected from each replicate. Hemolymph (0.5 mL) was collected from the foot base of the third leg by a 1.0 mL sterile syringe, then placed in a 1.5 mL centrifuge tube and mixed with an equal volume of anticoagulant solution (sodium citrate 13.2 g/L, citric acid 4.8 g/L, glucose 14.7 g/L). The mixture was immediately centrifuged at 4 ℃ and 4024 g for 20 minutes to obtain the supernatant, which was used to evaluate Acid phosphatase (ACP), alkaline phosphatase (AKP), lysozyme (LZM), total antioxidant capacity (T-AOC), Catalase (CAT), total superoxide dismutase (T-SOD), malondialdehyde content (MDA) and glutathione peroxidase (GSH-PX). Then, the hepatopancreas and intestine were collected into a 5 mL centrifuge tube pre-filled with 3 mL Bouin’s fixative for the production of intestinal and hepatopancreas tissue slices, respectively. Additionally, for each tank, two other crabs were randomly selected. Prior to dissection, the external surface of each crab was wiped with 75% ethanol to prevent external contamination. The entire intestine and hepatopancreas were dissected aseptically, frozen immediately in liquid nitrogen, and stored at -80 ℃ for the determination of intestinal microbial and digestive enzyme activities, respectively.
2.3.1. GROWTH PERFORMANCE
Survival, weight gain (WG), feed conversion ratio (FCR) and feed intake (FI) were calculated by combining the initial weight, final weight, the number of crab and the amount of diets consumed.
WG (%) = 100 × [(final weight (g) - initial weight (g)) / initial weight (g)].
Survival (%) = 100 × (final crab number ∕ initial crab number).
FCR = feed intake (g) / [final weight (g) - initial weight (g)].
FI (g/crab) = feed consumption (g) / [(final crab number + initial crab number)/2].
2.3.2. PROXIMATE COMPOSITION OF FEED AND CRAB
Moisture, crude protein, crude lipid, and ash of the experiment feed and crabs were measured following the method of AOAC.18 Moisture content was determined by drying the samples in an oven at 105 °C to a constant weight. Crude protein content was measured using an automatic Kjeldahl nitrogen analyzer (2300-Auto-analyzer, Foss Tecator, Sweden). Crude lipid content was assayed by the chloroform-methanol method. Crude ash content was determined after incineration in a muffle furnace at 550°C for 12 h.
2.3.3. SERUM BIOCHEMICAL INDICES
Serum biochemical indices, including acid phosphatase (ACP), alkaline phosphatase (AKP), lysozyme (LZM) and total antioxidant capacity (T-AOC), Catalase (CAT), total superoxide dismutase (T-SOD), malondialdehyde content (MDA), and glutathione peroxidase (GSH-PX) were measured according to the instructions of the reagent kit from Nanjing Jiancheng Biotechnology Research Institute, Nanjing, Jiangsu, China.
2.3.4. DIGESTIVE ENZYME ACTIVITY
Frozen hepatopancreas was mixed with 9 times saline (W:V=1:9), then homogenized and centrifuged at 4 ℃ for 15 min (1006 g). The supernatant was taken for the determination of digestive enzyme activity. The activities of protease, lipase and α-amylase were measured using their respective kits (A080-2-2, A054-2-1, C016-1-1) which from Jian Cheng Biochemical Company in Nanjing of China.
2.3.5. HEPATOPANCREAS AND INTESTINE HISTOLOGY
The preparation of hepatopancreas and intestinal tissue sections followed the methodology described in our previously published study.19 The intestines and hepatopancreas tissues were sequentially dehydrated with ethanol, cleared with xylene, and embedded in paraffin for sectioning. After H&E staining, the morphological characteristics were observed under an optical microscope, and relevant data were measured.
2.3.6. INTESTINAL MICROBIAL STUDY
The sequencing experiment was entrusted to Shanghai Majorbio Bio-pharm Technology Co., Ltd. (China) for next-generation sequencing (NGS) library preparation and Illumina Miseq sequencing. The specific method is as follows:
Total microbial genomic DNA was extracted from intestine using the FastPure Stool DNA Isolation Kit (MJYH, shanghai, China) according to manufacturer’s instructions. The quality and concentration of DNA were determined by 1.0% agarose gel electrophoresis and a NanoDrop® ND-2000 spectrophotometer (Thermo Scientific Inc., USA), and the DNA was stored at -80 ℃ prior to further use. The hypervariable region V3-V4 of the bacterial 16S rRNA gene were amplified with primer pairs 338F (5’-ACTCCTACGGGAGGCAGCAG-3’) and 806R(5’-GGACTACHVGGGTWTCTAAT-3’) by an T100 Thermal Cycler (BIO-RAD, USA). The PCR reaction mixture including 4 μL 5 × Fast Pfu buffer, 2 μL 2.5 mM dNTPs, 0.8 μL each primer (5 μM), 0.4 μL Fast Pfu polymerase, 10 ng of template DNA, and ddH2O to a final volume of 20 µL. PCR amplification cycling conditions were as follows: initial denaturation at 95 ℃ for 3 min, followed by 27 cycles of denaturing at 95 ℃ for 30 s, annealing at 55 ℃ for 30 s, and extension at 72 ℃ for 45 s, and a single extension at 72 ℃ for 10 min, and ended at 4 ℃. All samples were amplified in triplicate. The PCR product was extracted from 2% agarose gel and purified. Then quantified using Synergy HTX (Biotek, USA).
Purified amplicons were pooled in equimolar amounts and paired-end sequenced on an Illumina NextSeq 2000 PE300 platform (Illumina, San Diego,USA) according to the standard protocols by Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China). After demultiplexing, the resulting sequences were quality-filtered with fastp (v0.19.6) and merged with FLASH (v1.2.11). Then the high-quality sequences were denoised using the DADA2 plugin in the Qiime2 (version 2020.2) pipeline with recommended parameters, which obtains single-nucleotide resolution based on error profiles within samples. Taxonomic assignment of ASVs was performed using the Naive bayes consensus taxonomy classifier implemented in Qiime2 and the SILVA 16S rRNA database (v138.2). Bioinformatic analysis of the gut microbiota was carried out using the Majorbio Cloud platform (https://cloud.majorbio.com).
2.4. STATISTICAL ANALYSIS
All data were presented as mean ± standard deviation and analyzed using SPSS 22.0 statistical software. A one-way analysis of variance (ANOVA) and Turkey’s multiple range tests were used to determine the statistical significance among groups. Statistical significance was determined at P < 0.05. In addition, a follow-up trend analysis was performed using orthogonal polynomial contrasts to determine whether the significant effect was linear and/or quadratic.
3. RESULTS
3.1. GROWTH PERFORMANCE
As shown in Table 2, dietary supplementation of chitin had a significant quadratic effect on the FBW, WG, and FCR of E. sinensis (P < 0.05). With increasing chitin supplementation, FBW and WG initially increased, then decreased, while FCR showed the opposite trend. Compared with the C1 group, the C3 and C4 group resulted in a significantly higher FBW, WG and a significantly lower FCR (P < 0.05). No significant differences were observed in any growth indicators between the C2 and C5 groups and the C1 group (P > 0.05).
3.2. WHOLE-BODY PROXIMATE COMPOSITION
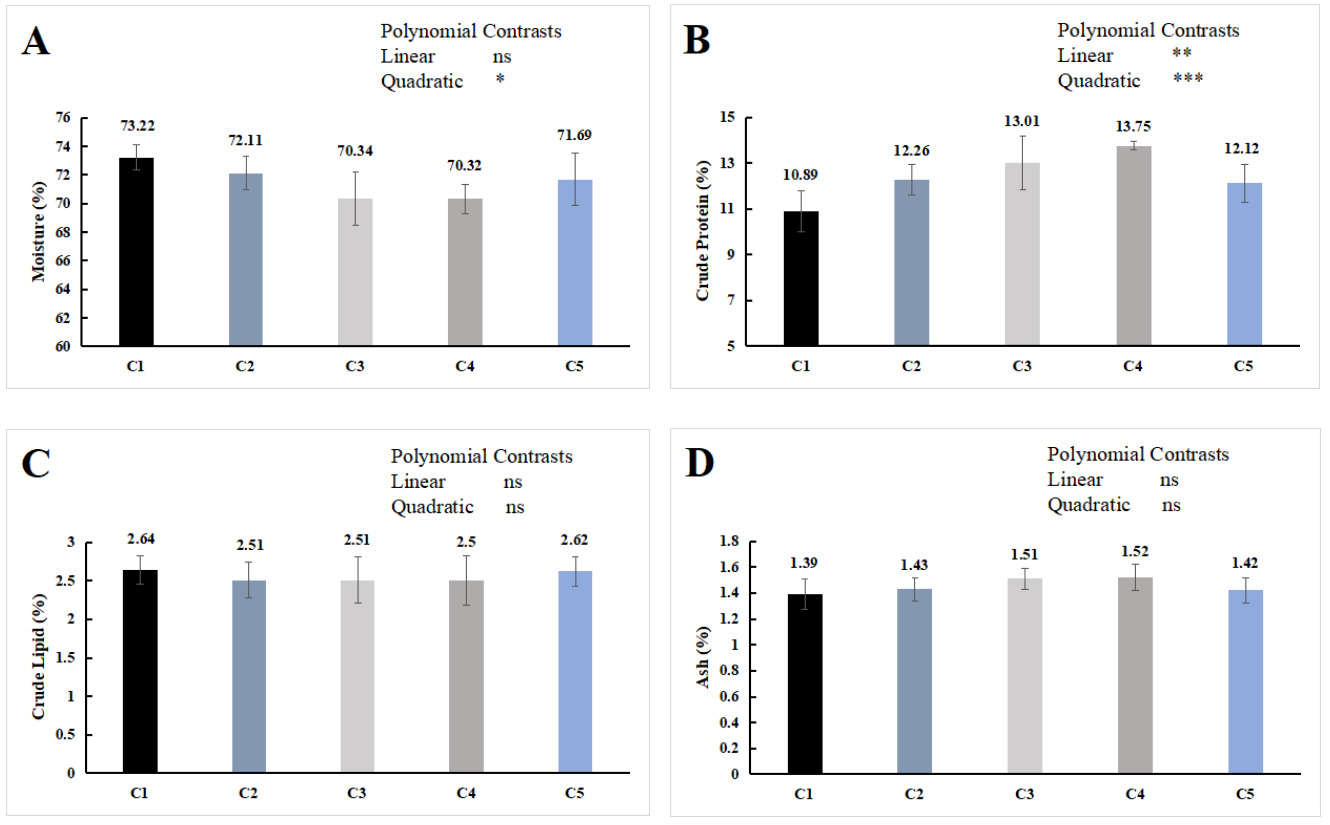
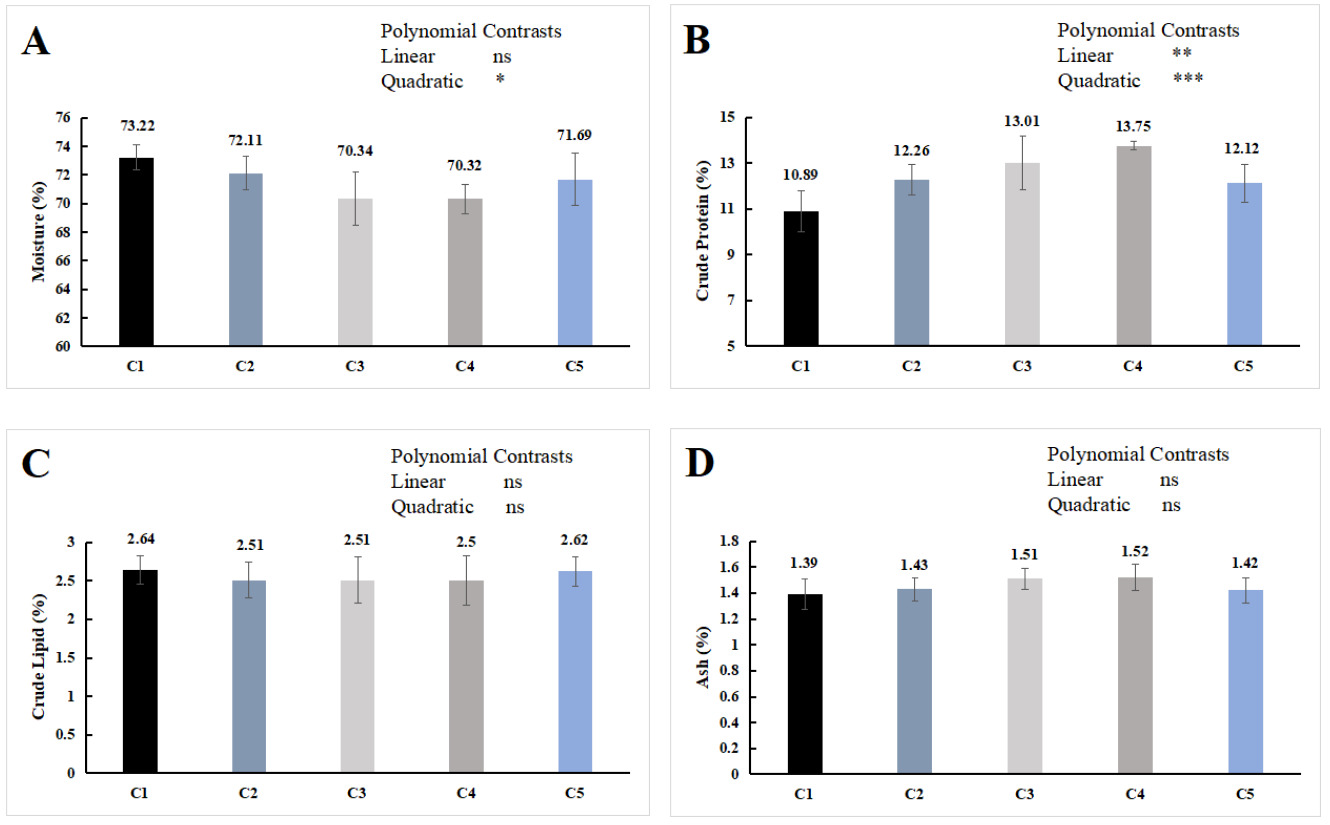
As shown in Figure 1, dietary chitin supplementation has no significant effect on the crude lipid and ash contents of the whole E. sinensis (P > 0.05) but exerted a significant quadratic effect on the moisture content (P < 0.05), and both significant linear and quadratic effects on the crude protein content (P < 0.05). With increasing chitin supplementation, the crude protein content initially increased and then decreased, while the moisture content showed the opposite trend. The crude protein content in the C3 and C4 groups was significantly higher than that in the C1 group (P < 0.05), and no significant difference were found in the proximate composition of the whole-body proximate composition among the groups of C2, C5 and C1 (P > 0.05).
3.3. DIGESTIVE ENZYMES ACTIVITIES IN HEPATOPANCREAS
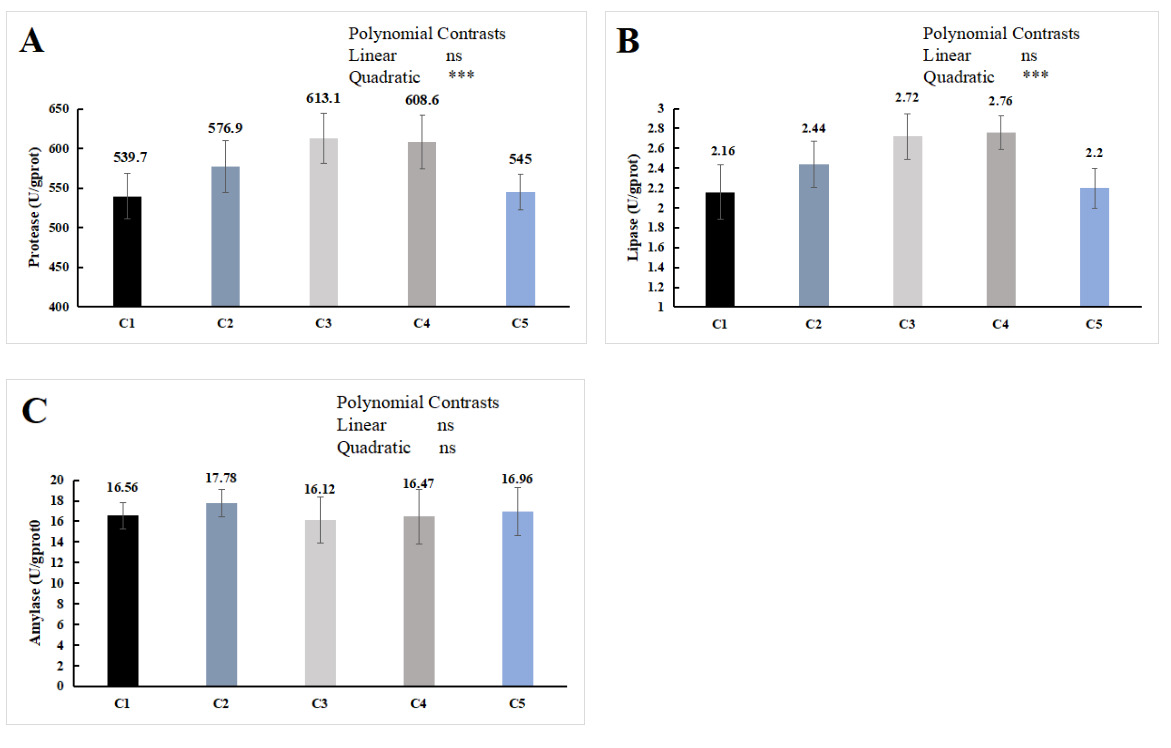
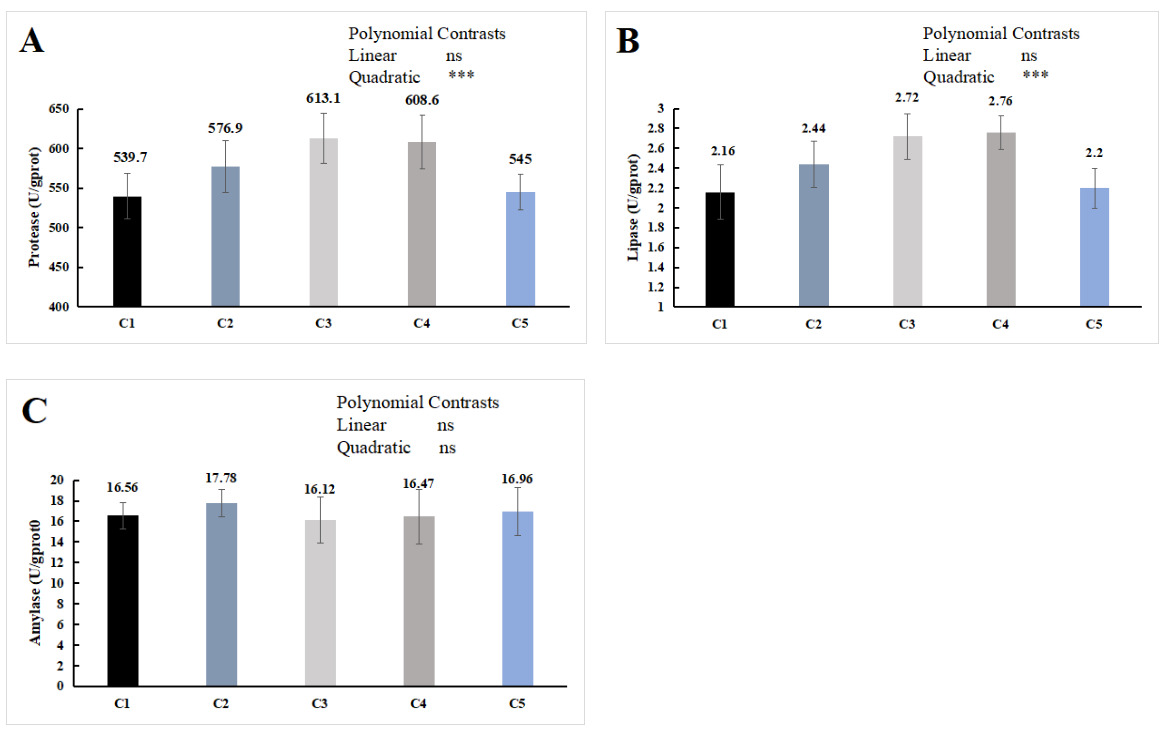
The orthogonal polynomial contrasts showed that the supplementation level of chitin quadratically increased the hepatopancreas protease and lipase activities, which were significantly higher in the C3 and C4 groups than in the C1 group (P < 0.05, Figure 2A, B). No significant difference was observed in the activity of the hepatopancreas amylase activity (P > 0.05, Figure 2C).
3.4. SERUM BIOCHEMICAL INDICES
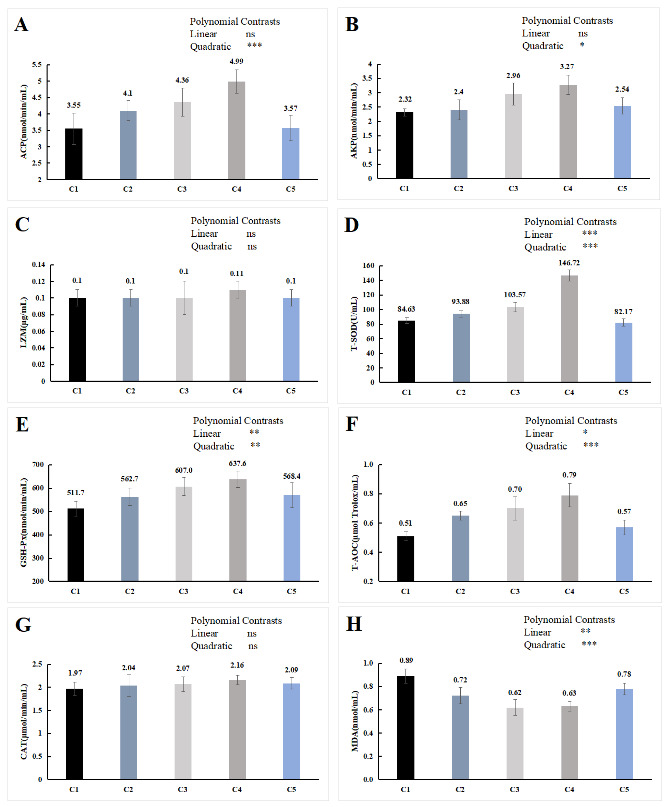
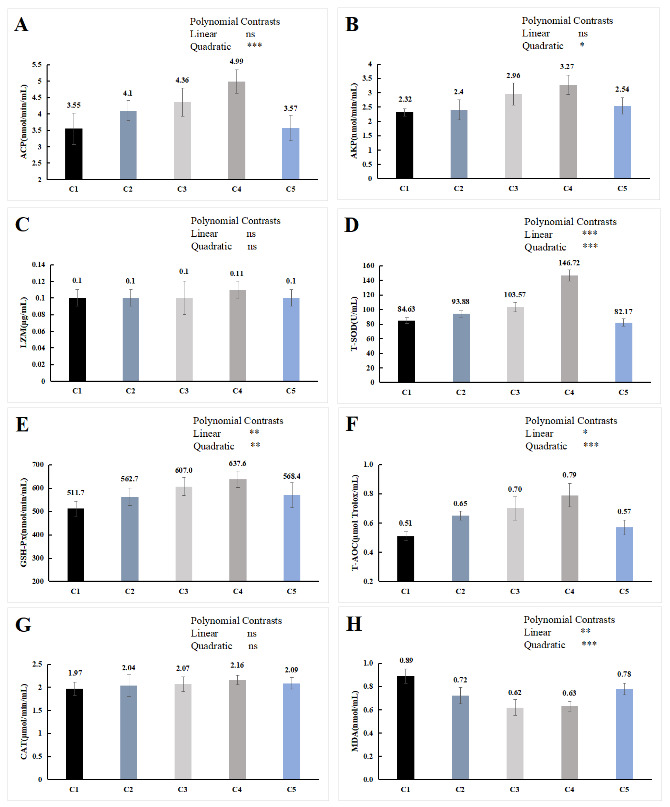
The orthogonal polynomial contrasts showed that the activity of ACP and AKP was quadratically increased with the increase in the supplementation level of chitin, which of the C4 group was significantly higher than the C1 group (P < 0.05, Figure 3A, B). The level of the supplementation of chitin has positive linear and quadratic effects on the activities of T-SOD, T-AOC and GSH-Px (P < 0.05, Figure 3D, E, F), and the content of MDA showed the opposite trend (P < 0.05, Figure 3H). Compared with the C1 group, T-SOD, T-AOC and GSH-Px activities in serum of C3 and C4 group were significantly higher (P < 0.05), the content of MDA was significantly lower (P < 0.05), while no significant difference in serum biochemical indicators between C5 group and C1 group (P > 0.05). In LZM and CAT, no significant difference was detected among all the treatments (P > 0.05, Figure 3C, G).
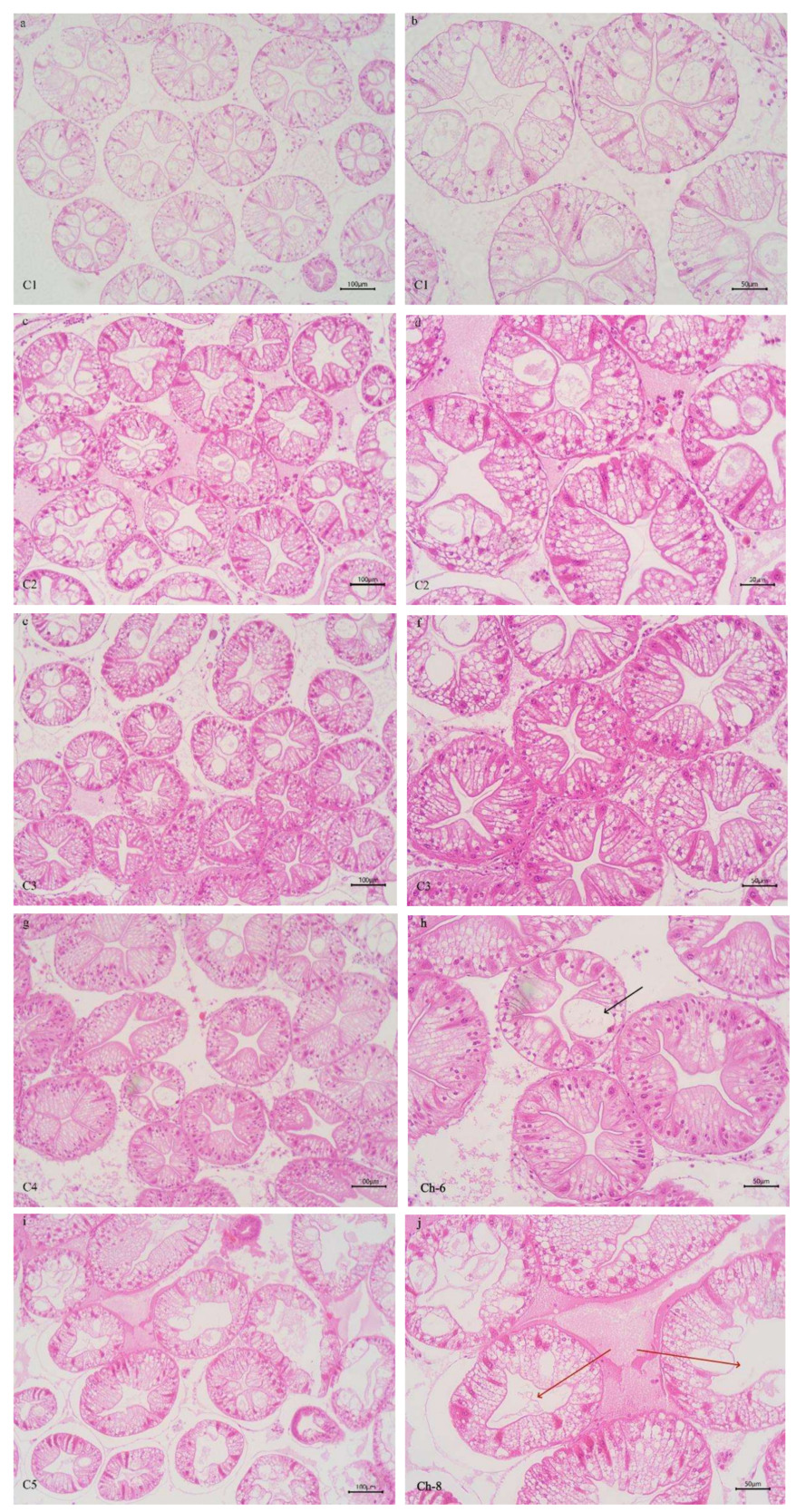
3.5. INTESTINAL HISTOLOGY
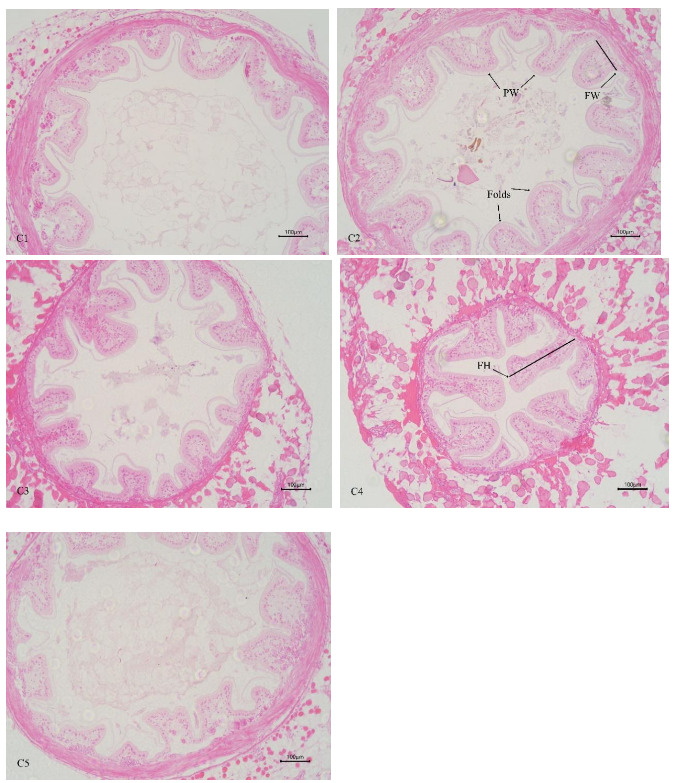
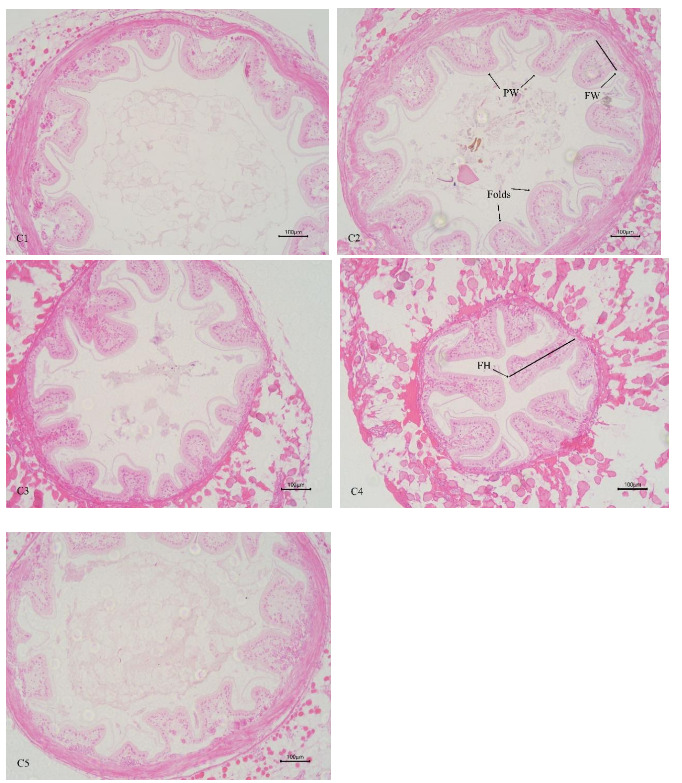
In Table 3 and Figure 4, the increasing level of chitin led to significant linear increase of mid-intestinal folds width, and quadratically increase of mid-intestinal folds number and height (P < 0.05). The folds number, height, and width of mid-intestinal in C1 group were significantly lower than those in C4 group (P < 0.05), and there was no significant difference between C2 and C5 groups (P > 0.05). There was no significant difference in mid-intestinal peritrophic membrane thickness among all the groups (P > 0.05).
3.6. Hepatopancreas histology
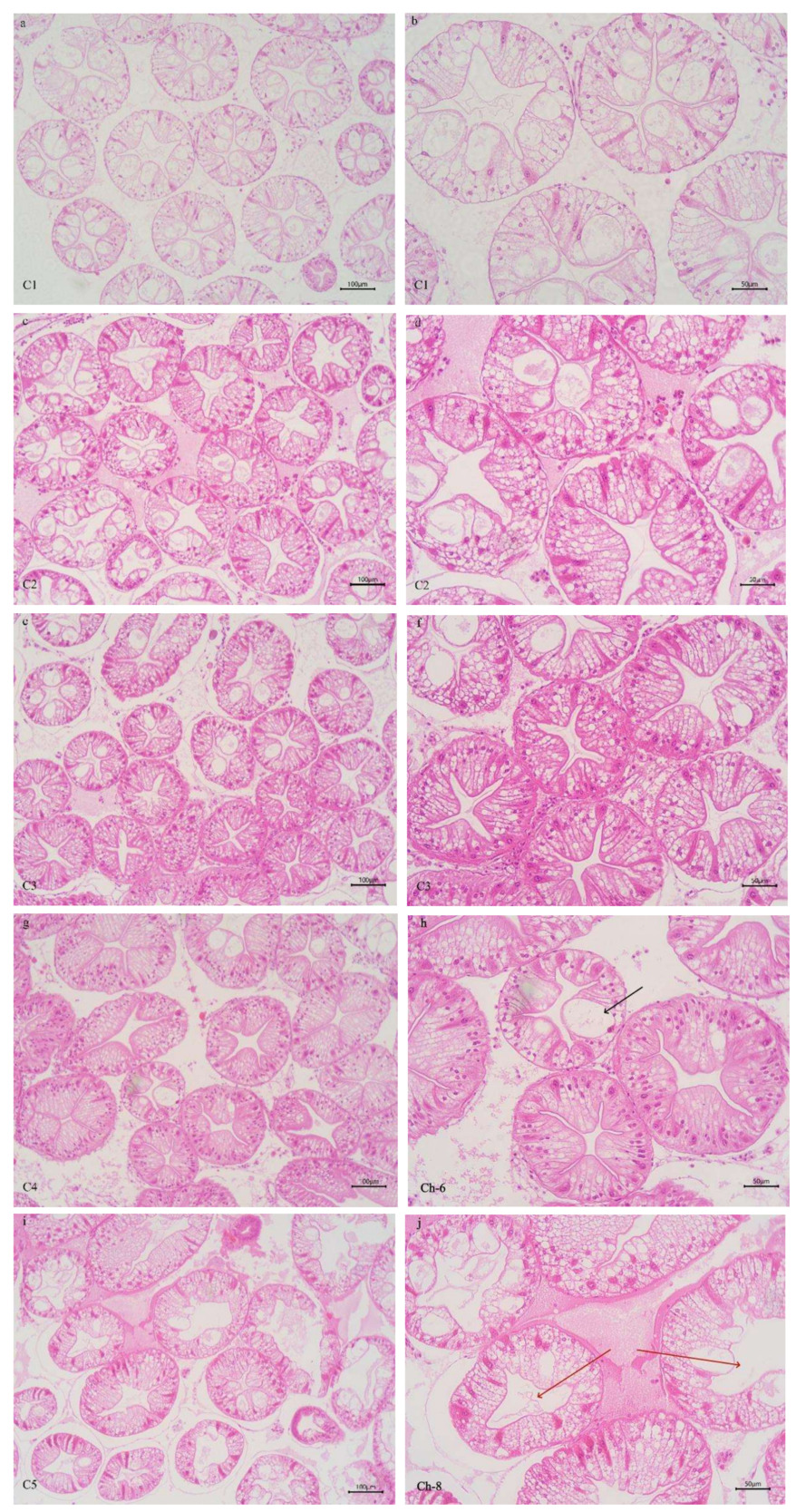
As shown in Figure 5, the intercellular junctions of the hepatopancreas tubular epithelial cells in the C3 and C4 groups were well-organized. The hepatopancreas tubules in these groups had a star-shaped and polygonal structure, with clear morphology, moderate lumen size and epithelial thickness. In contrast, in the C1 and C2 groups, morphological alterations were observed in some hepatopancreatic tubules of crab, including disappearance of the stellar structure, mild disorganization of cell arrangement, and cytoplasmic vacuolization of the tubular epithelial cells. In the C5 group, some hepatopancreatic tubules showed morphological changes, deformation of hepatopancreatic cells, severe structural damage, partial loss of intercellular boundaries, and cell fusion.
3.7. MICROBIAL STUDY IN INTESTINE USING 16 RRNA SEQUENCING
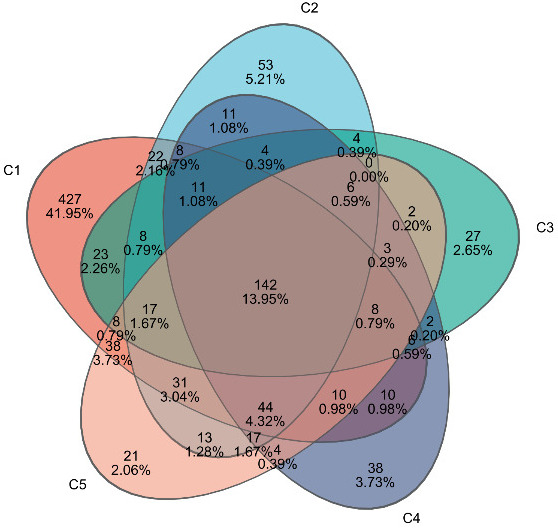
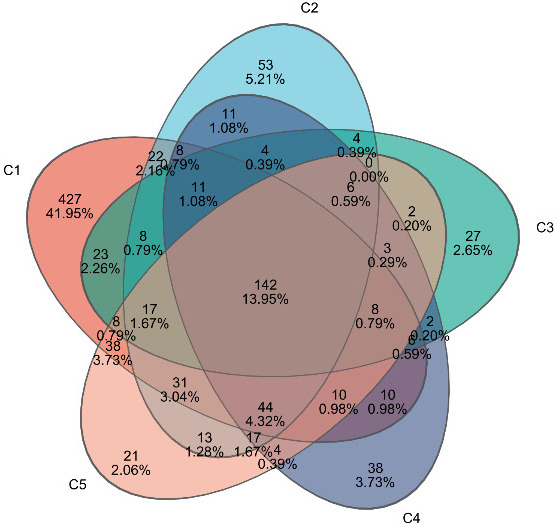
A total of 1,202,786 valid sequences were obtained with sequence lengths ranging from 421 to 440 bp, and the average sequence length was 423 bp. After cluster analysis of the obtained sequences, five groups obtained 813, 391, 279, 307, and 344 representative bacterial communities OTUs, respectively. The OTUs shared by the five treatment groups are 120, while the unique OTUs are 427, 53, 27, 38, and 21, respectively (Figure 6).
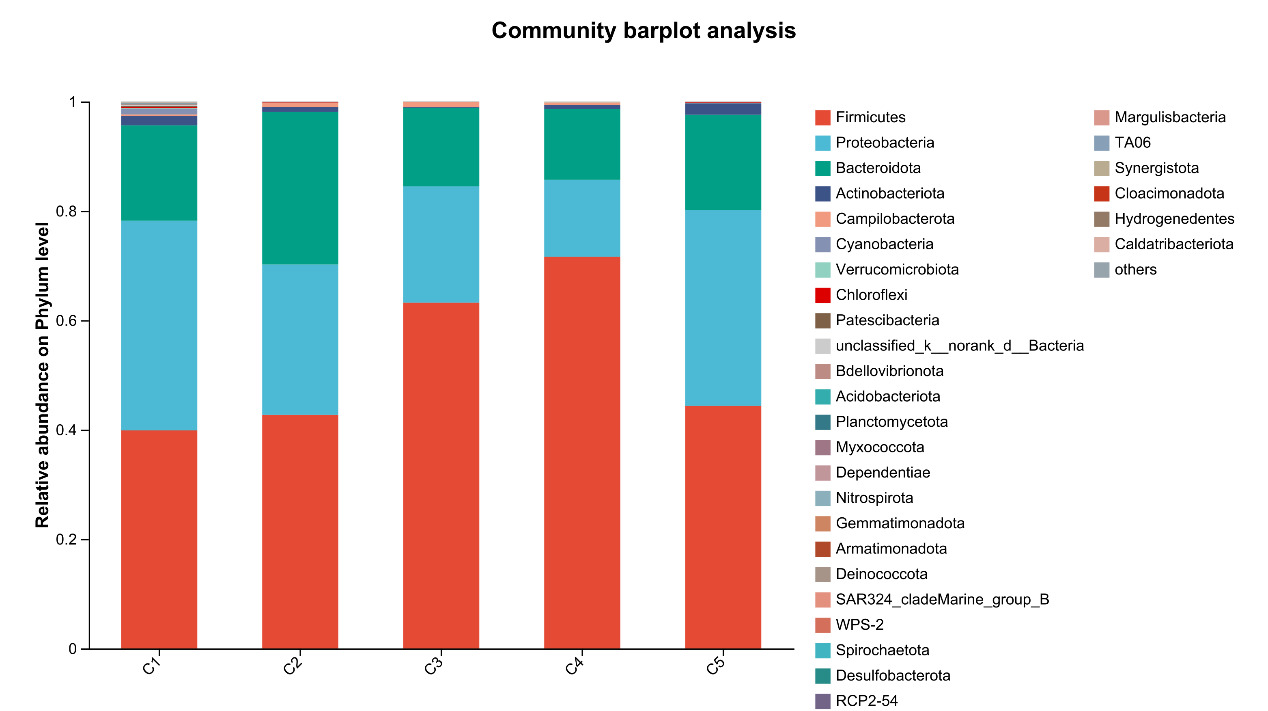
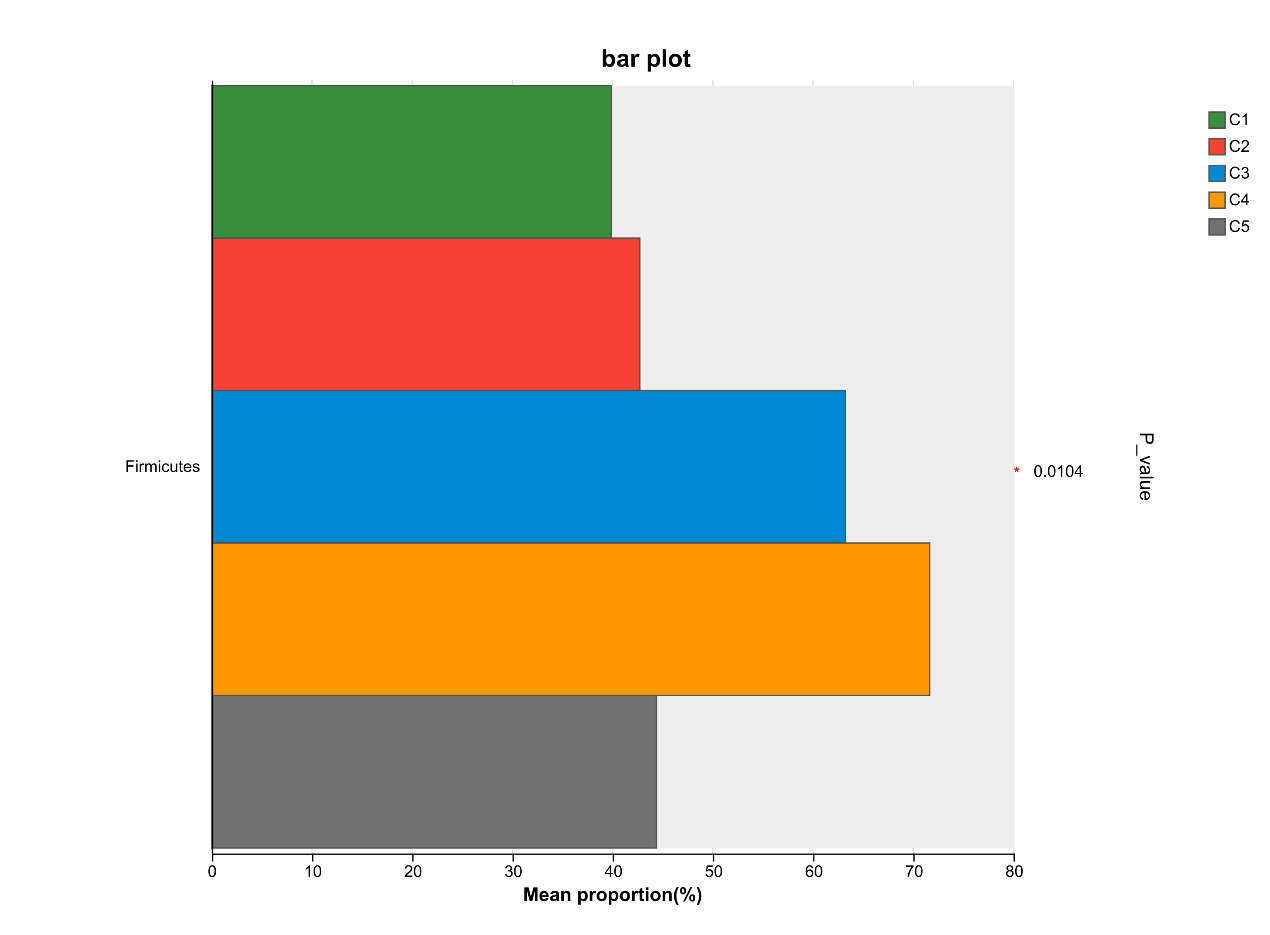
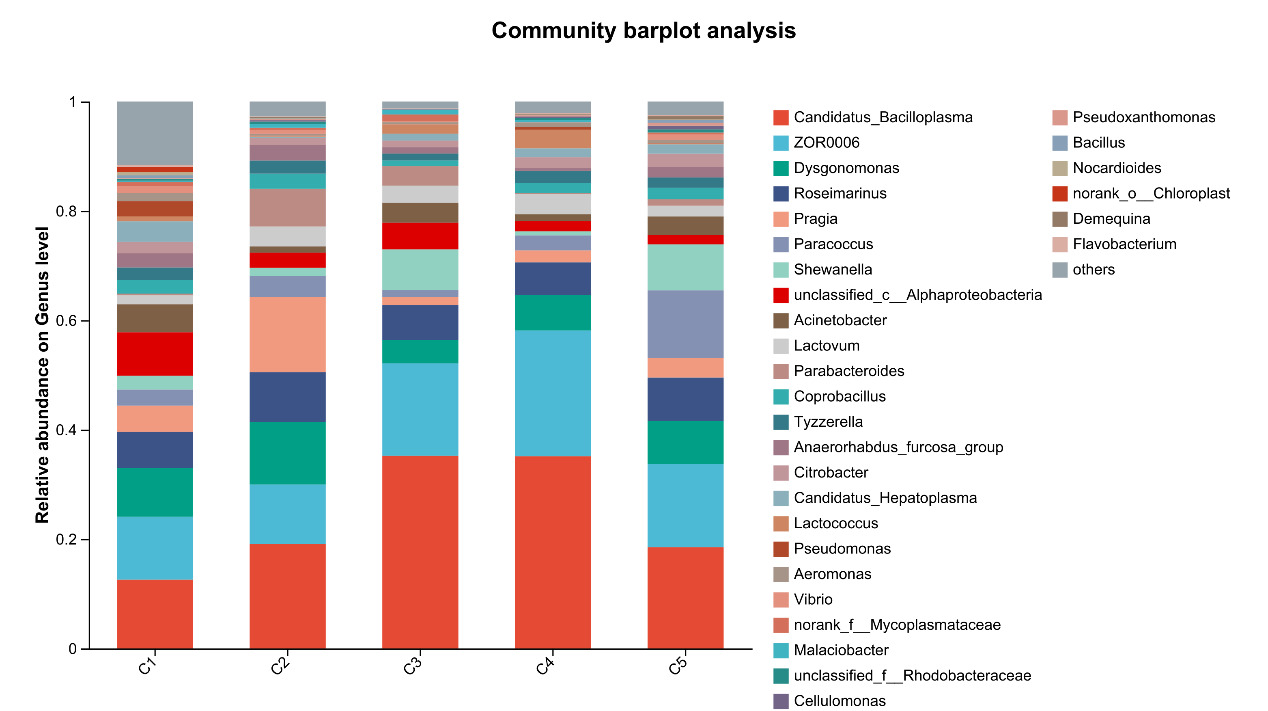
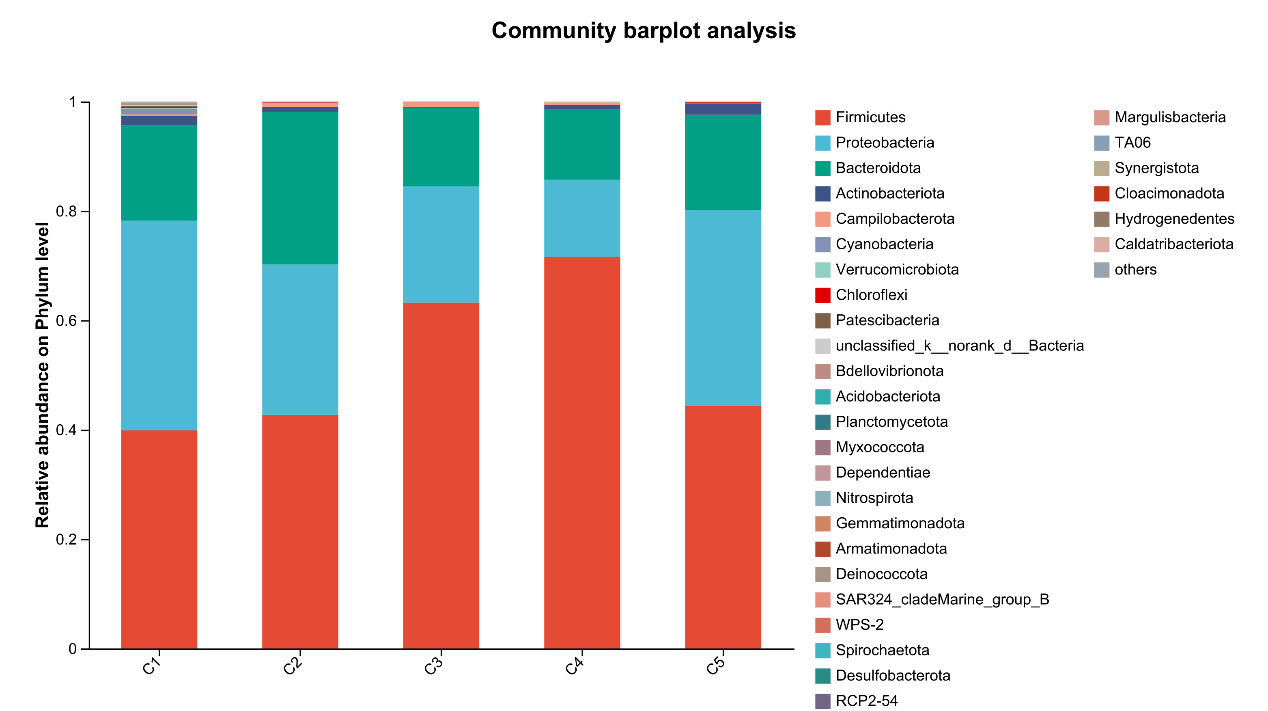
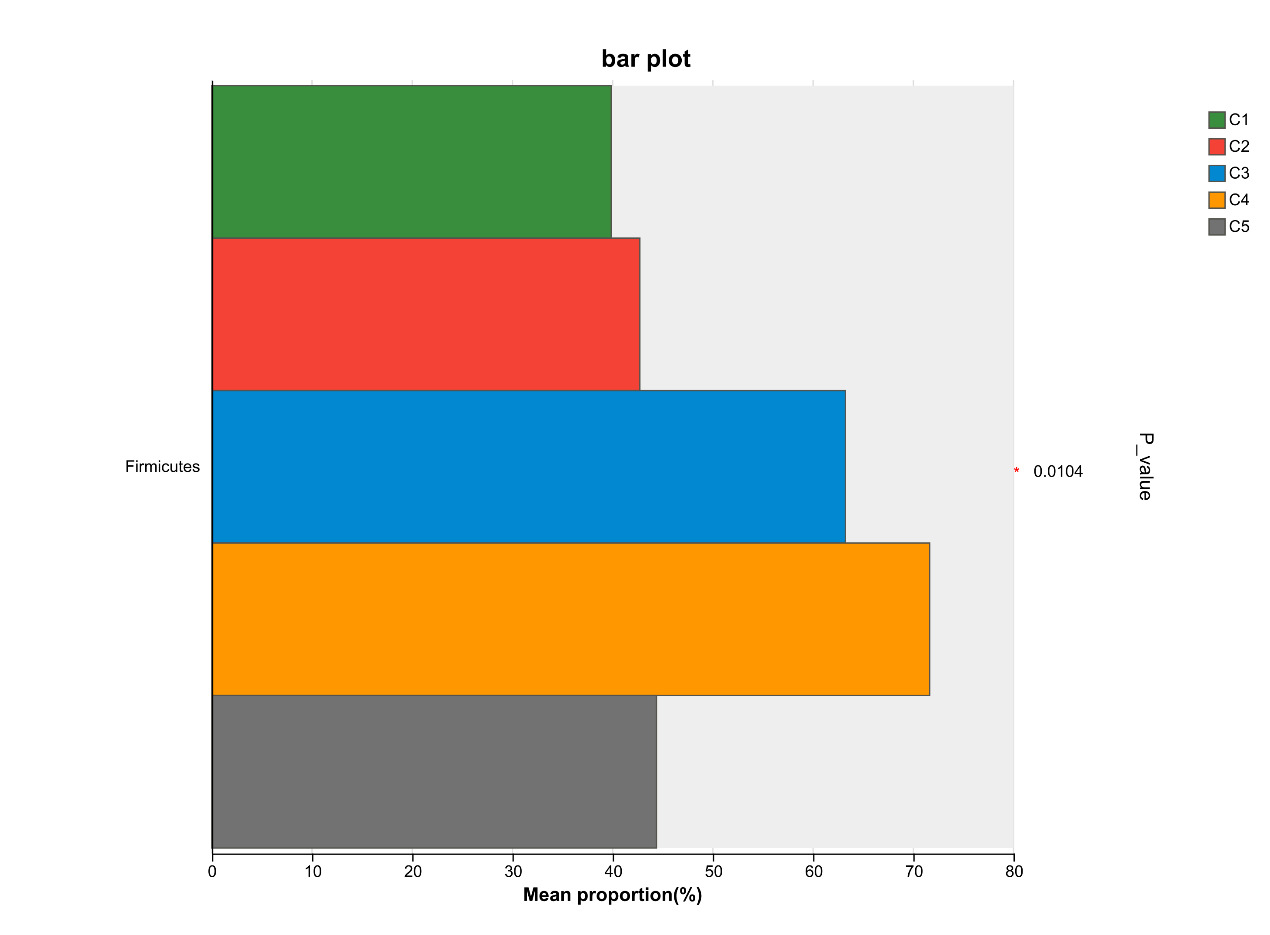
At the phylum level, Firmicutes, Proteobacteria, Bacteroidota, and Actinobacteriota are the most abundant bacteria in the intestinal microflora of E. sinensis, accounting for 97.37%, 99.00%, 99.02%, 99.38%, and 99.69%, respectively. The rest mainly include Campilobacterota, Cyanobacteria and Verrucomicrobiota. As shown in Figure 7, there were significant differences in the abundance of Firmicutes in the microbial composition of each treatment group (P < 0.05, Figure 8).
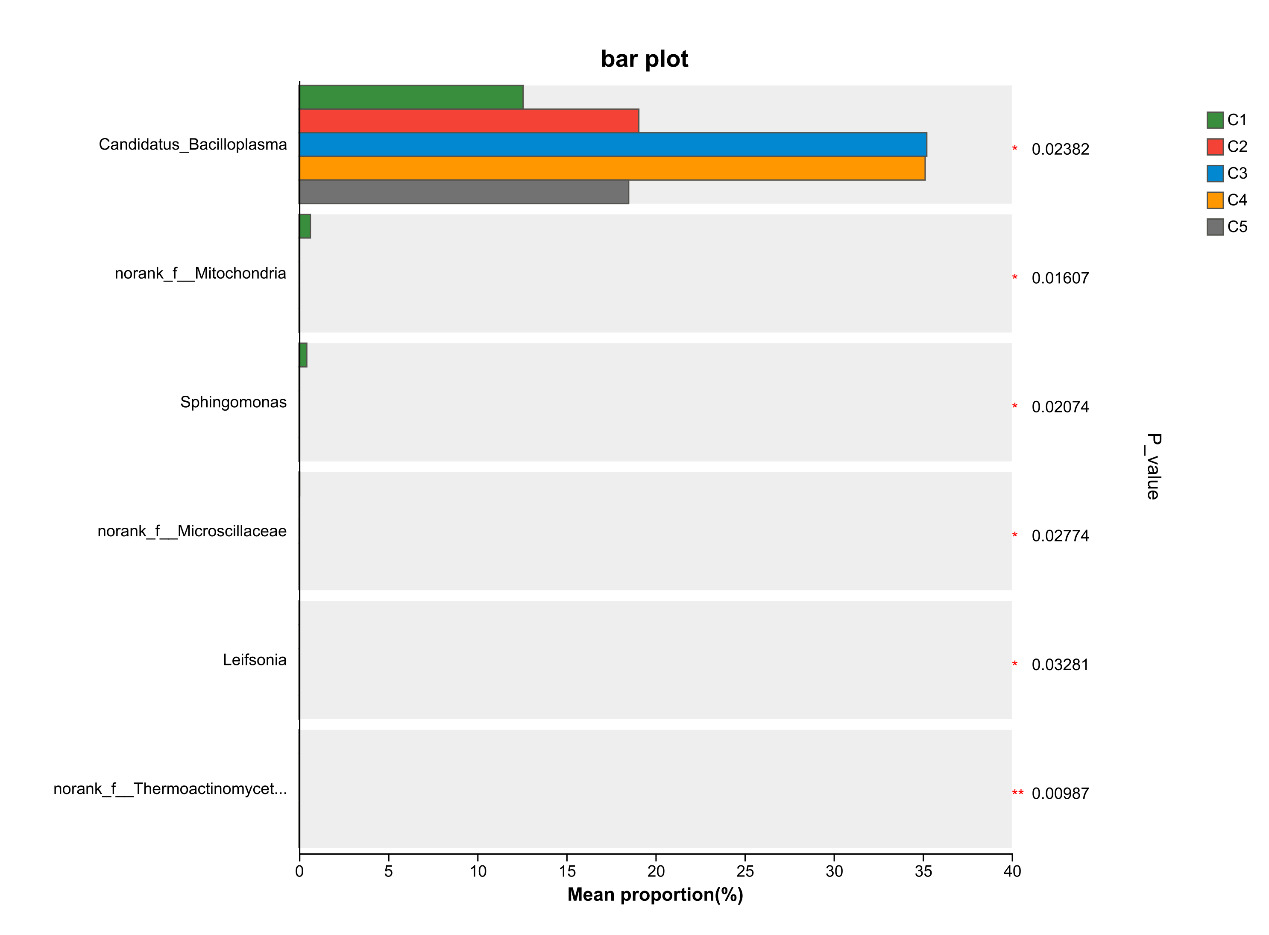
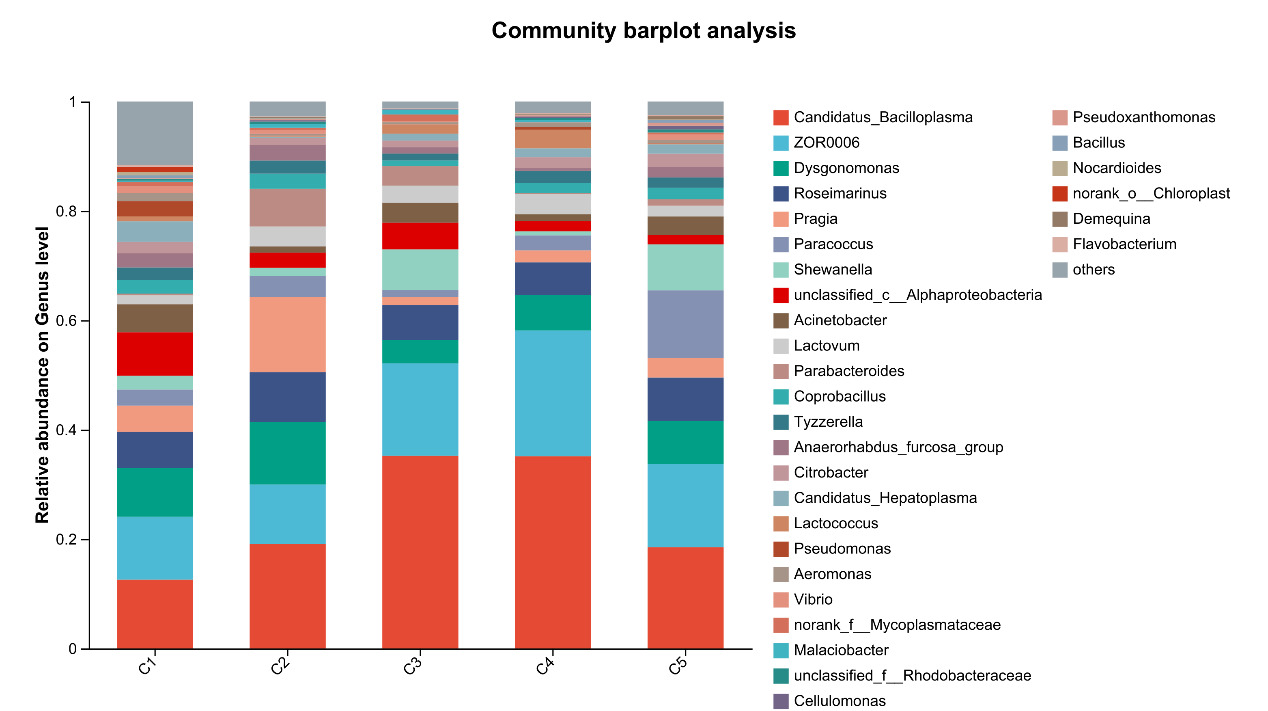
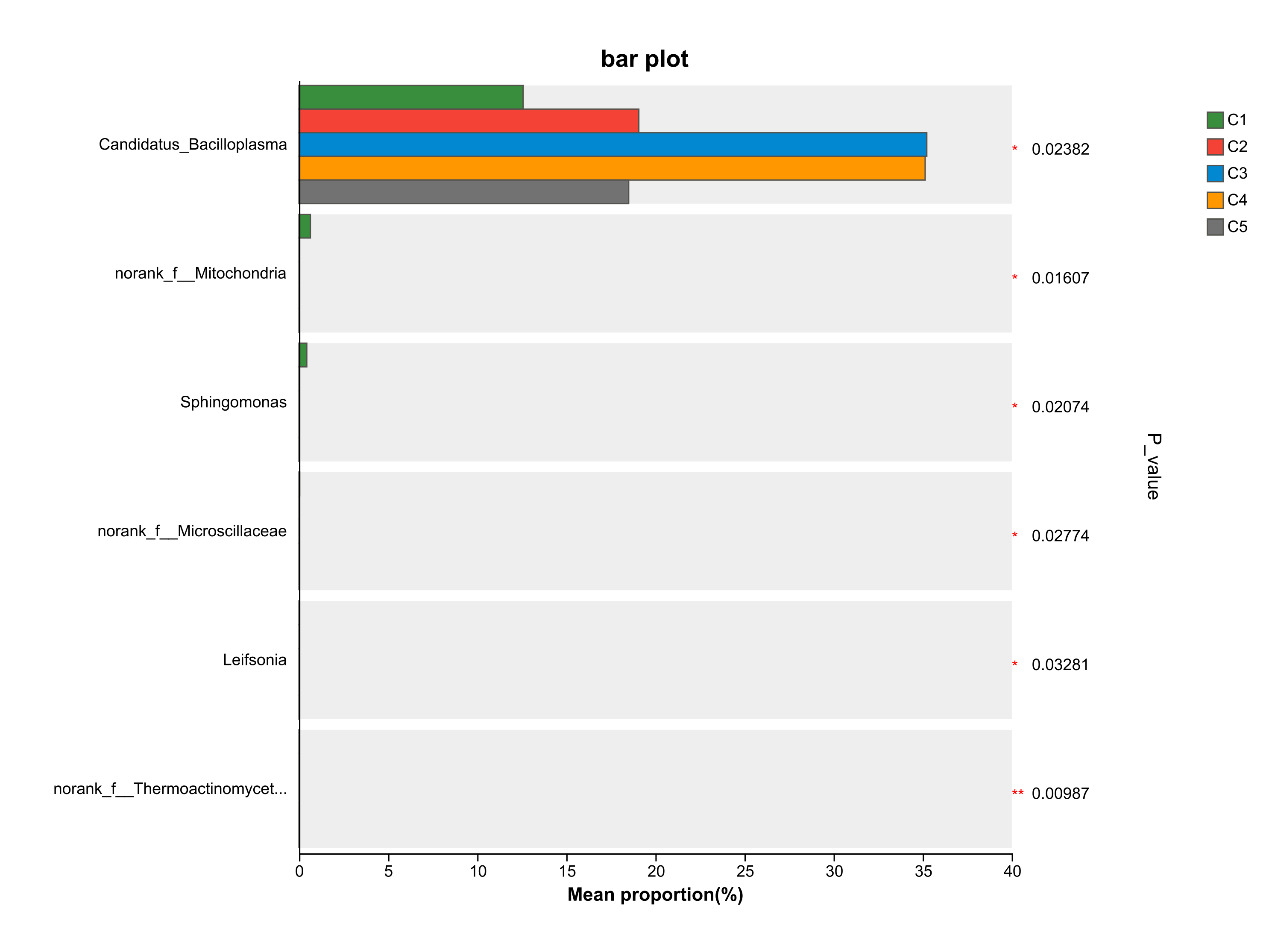
At the genus level, the dominant intestinal microflora included Candidatus_Bacilloplasma, ZOR0006, Dysgonomonas, Roseimarinus, Pragia, Paracoccus and Shewanella, the rest mainly include Acinetobacter, Lactovum, Parabacteroides and Coprobacillus (Figure 9). There were significant differences in the composition and abundance of gut microbiota among the treatment groups. The abundance of Candidatus_Bacilloplasm in C3 and C4 groups was significantly higher than that in other groups (P < 0.05). In addition, there were also significant differences in the abundance of norank_f_Mitochondria, Sphingomonas, norank_f_Microscillaceae, Leifsonia, norank_f_Thermoactinomycetaceae (P < 0.05, Figure 10).
4. DISCUSSION
In recent years, due to limited availability of fish meals and the rapid growth of the insect industry, insect protein has gained significant attention as a novel protein source. Multiple studies have confirmed the feasibility of replacing fish meals with insect protein.20,21 However, numerous studies have also reported that excessive substitution of fish meal with insect protein can lead to reduced growth performance in aquatic animals, a phenomenon largely attributed to the high chitin content in insect protein.22,23 As an essential nutrient, chitin must be degraded by chitinase in the digestive tract after ingestion to provide nutrients for the organism. So far, chitinase sequences have only been identified in a few fish species, and the presence of endogenous chitinase has not been reported in most economically important fish species. Consequently, high dietary chitin levels usually reduce nutrient utilization and growth performance in fish.24 As a typical crustacean, the Chinese mitten crab can secrete chitinase due to its habit of cannibalism or feeding on its own exoskeleton. Meanwhile, the intake of exogenous chitin can meet its energy requirements and support rapid exoskeleton regeneration. The present study found that adding 1.4%- 2.1% chitin to the diet significantly improved the growth performance of E. sinensis. Similar findings have been reported in black tiger prawn (Penaeus monodon)25 and giant freshwater prawn (Macrobrachium rosenbergii).26 The growth-promoting effect of chitin on crustaceans is closely associated with endogenous chitinase. Specifically, chitinase in the digestive tract catalyzes the hydrolysis of glycosidic bonds in dietary chitin, decomposing it into low-molecular-weight products such as chitooligosaccharides (COS) and N-acetylglucosamine. This process significantly enhances the bioavailability of chitin, facilitating efficient absorption by the organism and thereby promoting growth.27 However, with a further increase in dietary chitin content, the FBW of the C5 group was significantly lower than that of the C3 and C4 groups, while the FCR was significantly higher. Similar findings have been reported by Shiau et al.25 and Kumar et al.26 In their studies on Penaeus monodon and Macrobrachium rosenbergii, dietary chitin at 5% significantly improved weight gain and feed conversion efficiency, whereas chitin at 10% conversely reduced feed conversion efficiency. These results indicate that excessively high chitin content cannot be efficiently digested and absorbed by crustaceans, possibly due to endogenous chitinase activity. As dietary chitin content increases, the amount of chitin in the digestive system rises accordingly. The available chitinase may be insufficient to hydrolyze the excess chitin, leading to the formation of indigestible complexes with amino acids, which subsequently interfere with the digestion and absorption of other nutrients. In a related study, Yao et al.19 observed that replacing fish meal with high-chitin yellow mealworm meal in diets for E. sinensis resulted in significantly reduced growth performance and intestinal structural damage at the 100% replacement level, consistent with the findings of the present experiment.
The whole-body composition of Chinese mitten crab can be influenced by dietary formulation. In this experiment, dietary supplementation with 1.4%-2.1% chitin significantly reduced whole-body moisture content and significantly increased crude protein content compared with the control group (P < 0.05) (Figure 1). Similar findings have been reported in Pacific white shrimp (Litopenaeus vannamei) with 0.4% chitin supplementation28 and in giant freshwater prawn (Macrobrachium rosenbergii) with 5% chitin supplementation.26
Similar to cellulose, chitin has the potential to act as a prebiotic in aquatic animal feeds. It can promote the proliferation of beneficial intestinal bacteria, enhance intestinal microbial diversity, and stimulate intestinal fermentation to produce short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate. These mechanisms contribute to improved intestinal health and enhanced nutrient absorption and utilization.29 Furthermore, as chitin is hydrolyzed by chitinase into low-molecular-weight oligosaccharides (e.g., chitooligosaccharides), its bioavailability may be enhanced, allowing for more efficient absorption in the digestive tract.24 In addition, oligosaccharides have been shown to possess prebiotic properties, and dietary supplementation with oligosaccharides can enhance protein absorption.30 However, when the dietary chitin content is excessively high, the endogenous chitinase levels in Chinese mitten crab are insufficient to fully degrade the ingested chitin. The excess chitin binds to amino acids, forming indigestible complexes that interfere with the absorption of other nutrients. Consequently, when the chitin supplementation level exceeded 2.1% in this experiment, the whole-body protein content of the crab decreased instead of increasing (Figure 1). A similar trend was observed in intestinal histomorphology: the number and height of intestinal folds in the C5 group were significantly reduced compared with those in the C4 group and did not differ significantly from those in the C1 group (Table 3). These results indicate that an appropriate amount of dietary chitin can increase the whole-body protein content in Chinese mitten crab, whereas excessive chitin produces adverse effects.
As a typical crustacean species, E. sinensis lacks an adaptive immune system and relies entirely on innate immunity to defend against pathogens. The hepatopancreas serves as a vital immune organ in the innate immune system of E. sinensis, with key immune enzymes, including acid phosphatase (ACP), alkaline phosphatase (AKP), and lysozyme (LZM), widely recognized as critical indicators of its immunocompetence.31 In the present study, the activities of ACP and AKP in the hepatopancreas of the C4 group were significantly higher than those in the C1 group. Several studies have also demonstrated that dietary supplementation of chitin can significantly enhance the immune response, disease resistance, and survival rate of aquatic animals.6,32 Kumar et al.33 reported that the inclusion of 0.75% and 1% chitin in the diet significantly improved the immune responses of Macrobrachium rosenbergii, including prophenoloxidase (proPO) activity and respiratory burst (RB) of hemocytes, as well as improved resistance against white muscle disease (WTD). In a study on Litopenaeus vannamei conducted by Cheng et al.6 the results showed that dietary supplementation with 0.4% chitin could significantly upregulate the expression of antimicrobial protein genes, including pen3a, crus, and alf2, thereby enhancing their immune capacity. Chitin can regulate the immune function of aquatic animals through interactions with its receptors. Additionally, it can be degraded into chitosan in the intestinal environment, which further modulates the intestinal microbial community and protects the organism.34 However, the optimal addition of chitin varies among species due to differences in digestive and metabolic capacity. The results of the present study indicate that a dietary chitin level of 2.1% significantly enhanced the immune response of E. sinensis. Nevertheless, an excessively high supplementation level exerts the opposite effect, which may be attributed to the fact that crabs have difficulty digesting and absorbing chitin.
During metabolic processes, organisms produce a large amount of reactive oxygen species (ROS). An imbalance between ROS production and elimination can lead to oxidative stress in aquatic animals.35 In the process of resisting oxidative damage, aquatic animals have developed an antioxidant system, which includes antioxidant substances and enzymes such as glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), catalase (CAT), total antioxidant capacity (T-AOC), and malondialdehyde (MDA).36 In the present study, it was observed that the activities of hepatopancreatic T-SOD, GSH-Px and T-AOC in the C3 and C4 groups were significantly higher than those in the C1 group, while the MDA content was significantly lower than that in the C1 group. Consistently, Niu et al.37 reported that dietary supplementation with 0.4% chitin in Penaeus monodon significantly increased the hepatopancreatic GSH-Px activity and T-AOC, and decreased the MDA content compared with the control group. These effects may be attributed to chitosan oligosaccharide, a derivative of chitin. Chitosan oligosaccharide, one of the hydrolysis products of chitin in crustaceans, exhibits strong antioxidant activity. It reacts with high-energy free radicals via amino and hydroxyl groups on its pyranose ring, facilitating the formation of stable molecular complexes.38 Additionally, it has been shown to alleviate oxidative stress by activating the inflammatory pathway involving nuclear factor erythroid 2-related factor 2 (Nrf2), or by inhibiting signaling pathways such as nuclear factor kappa-B (NF-κB) and mitogen-activated protein kinase (MAPK).39 A study by Niu et al.37 demonstrated that dietary supplementation with 4 g/kg chitosan oligosaccharide enhanced the activities of GSH-Px and T-AOC in the hepatopancreas of Penaeus monodon, while reducing MDA content. Similar findings were reported by Guan et al.40 in Nile tilapia (Oreochromis niloticus) and by Fu et al.41 in Chinese soft-shelled turtle (Pelodiscus sinensis). Therefore, appropriate dietary chitin supplementation can enhance the antioxidant capacity of aquatic animals. However, optimal supplementation levels vary among aquatic animal species, warranting further investigation.
In crustaceans, the intestine harbors diverse microbial communities that not only participate in host digestion, absorption, and nutrient exchange but also play crucial roles in maintaining host health and regulating many essential physiological functions.42 The homeostasis of the intestinal microbiota in E. sinensis is vital for its metabolism and immunity. A healthy gut microbial community can produce beneficial metabolites and protect the host from pathogenic microorganisms. In contrast, an imbalanced microbial community may lead to the production of harmful metabolites, compromise host immunity, and increase susceptibility to diseases.43 In the present study, the dominant intestinal microbiota of E. sinensis across all treatment groups were identified as Firmicutes, Proteobacteria, Bacteroidota, and Actinobacteriota. Collectively, these four phyla accounted for 97.37%, 99.00%, 99.02%, 99.38%, and 99.69% of the intestinal microbiota, respectively. Among them, significant differences were observed in the abundance of Firmicutes, with the C4 group showing a significant increase compared to the C1 group (P < 0.05). Firmicutes are involved in the transformation of complex macromolecular organic substances such as proteins and lipids during host growth, providing nutrients and energy to the organism.44 They are considered beneficial intestinal bacteria and have been shown to positively influence growth performance, immunity, digestive capacity, and disease resistance in aquatic animals.45 Similarly, at the genus level, the abundance of Candidatus_Bacilloplasma in the C4 group was significantly higher than that in the C1 group (P < 0.05). Candidatus_Bacilloplasma is a member of the indigenous intestinal microbiota, serving as a symbiotic microorganism involved in the digestive system of crustaceans.46 Previous studies have reported that Candidatus_Bacilloplasma is closely associated with the digestive function of E. sinensis and is considered beneficial to the host.47 Yang et al.48 also observed a significantly higher relative abundance of Candidatus_Bacilloplasma in the intestine of healthy crabs compared to diseased individuals. The findings of the present study indicate that dietary supplementation with 2.1% chitin has a positive impact on the crab’s intestinal microbiota, consistent with the observed intestinal histomorphology. The intestinal surface of E. sinensis is equipped with well-developed intestinal mucosal folds, which primarily function to increase intestinal surface area, thereby enhancing the absorptive, contact, and secretory areas of the intestine, ultimately facilitating digestion and absorption.49 In the present study, the number, height, and width of intestinal folds in the C4 group were significantly higher than those in the C1 group (P < 0.05) (Table 3, Figure 4). This finding indicates that an appropriate amount of chitin can promote the healthy development of the crab intestine. Chitin or its derivative chitosan in the feed possesses prebiotic properties. They can enhance the intestine bacterial diversity by promoting the proliferation of beneficial bacteria and chitin-degrading bacteria.33 This process further stimulates intestinal fermentation to produce essential short-chain fatty acids (SCFAs), among which acetic acid, propionic acid, and butyric acid are the main end products of bacterial fermentation. Such a mechanism contributes to improve the intestinal health and condition of aquatic organisms.29 However, when chitin was supplemented in excess, its positive effects on intestinal microbiota and tissue structure no longer manifested. No significant differences were observed in all intestinal indicators of the C5 group compared with the C1 group (Table 3, Figure 4). Although no adverse effects were detected in the present study with a 2.8% chitin supplementation level, the number and height of intestinal folds exhibited a significant quadratic effect with increasing chitin addition. Thus, a further increase in chitin content may potentially impair intestinal health in crabs. Yao et al.50 replaced 75% of fish meal in E. sinensis feed with black soldier fly meal, which has a high chitin content, resulting in a significant increase in the abundance of the harmful bacterium Shewanella, a significant decrease in the number and height of intestinal folds, and damage to fold integrity. Similar results were also observed in the application of yellow mealworm meal in the diet of E. sinensis.19 It can be concluded that supplementation of an appropriate amount of chitin in feed exerts a positive effect on intestinal health, whereas excessive inclusion may have detrimental effects.
Due to the absence of a gallbladder, the hepatopancreas of crustaceans serves as a critical organ for digestion, immunity, detoxification, and lipid storage, with functions analogous to those of the liver and pancreas in mammals. In healthy crabs, the hepatopancreas is composed of numerous hepatopancreatic tubules, which are the basic structural and functional units and contain central lumen. Each tubule consists of a central lumen surrounded by a single layer of columnar epithelial cells resting on a basement membrane. These cells feature neatly arranged nuclei and distinct intercellular boundaries.51 In the present study, hematoxylin-eosin (H&E) staining revealed that the hepatopancreatic structure of crabs fed diets supplemented with 1.4%-2.1% chitin was significantly improved compared with the control group. The tubules displayed intact cellular architecture, distinct star-shaped lumens, and reduced vacuolization (Figure 5). These improvements may be attributed to the fact that the hepatopancreas is the primary site of chitinase secretion.
Chitinases [1,4-(N-acetyl-β-D-glucosaminide) glycanohydrolase, EC 3.2.1.14)] are a class of glycosyl hydrolases and constitute a multi-gene enzyme family that degrades chitin by hydrolyzing the β-1,4-glycosidic bonds of chitin polymers or oligomers.52 To date, chitinase genes have been found to be highly expressed in the hepatopancreas of various crustaceans, including Macrobrachium nipponense,53 Penaeus monodon,54 Litopenaeus vannamei55 and E. sinensis.52 When crabs ingest an appropriate amount of chitin, it stimulates the secretion of chitinase from the hepatopancreas, which degrades chitin into small-molecule compounds. One of the degradation products, chitosan oligosaccharide, has been demonstrated to improve hepatopancreatic health.56 The hepatopancreas is also the primary site of secretion for other digestive enzymes. In the present study, the activities of hepatopancreatic protease and lipase in the C3 and C4 groups were significantly higher than those in the C1 group (Figure 2), which is consistent with the findings of hepatopancreatic histological structure observations, indicating that appropriate dietary chitin supplementation can improve the hepatopancreatic health of crabs and promote digestion and absorption. However, excessive chitin inclusion caused structural damage to the hepatopancreas. As observed in this study, the morphology of some hepatic tubules in the C5 group was altered, hepatopancreatic cells were deformed, and cell structures were severely damaged. Additionally, the activities of hepatopancreatic protease and lipase in the C5 group were significantly lower than those in the C3 group (Figure 2). These findings demonstrate that dietary chitin supplementation at 1.4-2.1% improves hepatopancreatic health in Chinese mitten crabs, whereas inclusion levels exceeding 2.1% cause structural impairment and reduce digestive enzyme secretion.
5. CONCLUSION
In summary, dietary supplementation with 1.4-2.1% chitin improved the growth performance and immune capacity of E. sinensis, whereas excessive inclusion levels induced negative effects.
ACKNOWLEDGMENTS
This work was financially supported by Jiangsu Agri-animal Husbandry Vocational College school level research project (NSF2025ZR08) and Jiangsu Haorun Biological Industry Group Co., Ltd. (S20240619).
AUTHORS’ CONTRIBUTION - CRediT
Methodology: Chunyan Zhang, Ningyang Sun, Guoan Hua, Sizhe Zhao.
Formal Analysis: Chunyan Zhang, Ningyang Sun, Yang Yang, Sitong Zhao, Dongbo He. Investigation: Chunyan Zhang, Ningyang Sun. Writing – original draft: Yang Yang, Sitong Zhao, Guoan Hua, Sizhe Zhao. Writing – review & editing: Chunyan Zhang, Ningyang Sun. Conceptualization: Chunyan Zhang, Ningyang Sun. Funding acquisition: Chunyan Zhang, Ningyang Sun, Quan Wang, Wenxiang Yao. Project administration: Chunyan Zhang, Ningyang Sun, Dongbo He, Quan Wang, Wenxiang Yao.
ANIMAL ETHICS STATEMENT
All animal care and use procedures were approved by the Institutional Animal Care and Use Committee of Jiangsu Agri-animal Husbandry Vocational College (Permit number: jsahvc-2023-17), and all authors clearly indicated that such guidelines have been followed.
DECLARATION OF COMPETING INTEREST
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of this article.
DATA AVAILABILITY
Data will be made available on request.