Introduction
The black tiger shrimp (Penaeus monodon), also known as the giant tiger prawn, is one of the most economically important marine shrimp species worldwide and is widely distributed in tropical and subtropical coastal regions. Owing to its large body size, superior flesh quality, and high market value, it is highly favored in premium seafood markets. P. monodon exhibits strong environmental adaptability and can be cultured in saline and brackish waters, making it suitable for pond culture, intensive tank systems, and recirculating aquaculture systems. According to the China Statistical Yearbook (2023), domestic production of P. monodon reached 128,420 tons. Despite continued production growth, domestic supply remains insufficient to meet market demand, and large quantities are still imported from Southeast Asian countries. This situation highlights the need for more efficient intensive culture strategies. During the transition toward intensive culture, stocking density has become a key management factor affecting shrimp performance and culture stability. Excessively high stocking density and feeding rates can alter key physicochemical parameters of the culture environment, including dissolved oxygen, ammonia nitrogen, nitrite, and pH, thereby increasing environmental pressure on cultured shrimp.1,2 High stocking density has also been reported to affect gut microbiota composition and immune status in shrimp.1 In addition, elevated nitrogen and phosphorus loads can impose direct physiological stress on shrimp, leading to growth inhibition, oxidative stress, altered antioxidant defense, and tissue damage in Litopenaeus vannamei.3 Previous studies in P. monodon have shown that elevated stocking density can reduce growth performance and survival in pond and nursery systems.
Although the effects of stocking density on growth performance and survival have already been documented in P. monodon,4–6 previous studies in this species have mainly focused on production traits rather than intestinal microbial responses under indoor culture conditions. In contrast, the intestinal microbiota of penaeid shrimp has been shown to vary with host developmental stage, diet, and culture-related conditions.7,8 Therefore, the key question in the present study was not whether stocking density affects P. monodon in general, but whether different stocking densities are associated with coordinated changes in growth performance, water quality, and gut microbial community structure under controlled indoor culture conditions.9 Based on this rationale, we hypothesized that increasing stocking density would reduce growth performance, increase environmental loading, and alter gut microbial diversity and community composition in P. monodon under indoor culture conditions.
Accordingly, this study aimed to evaluate the effects of different stocking densities on growth performance, water quality, and intestinal microbial communities of P. monodon under controlled indoor conditions.
Materials and Methods
2.1 Experimental Animals and Culture Conditions
The experiment was conducted at the Shenzhen Experimental Base of the South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences. A total of 1,050 healthy juvenile P. monodon was selected, with an average body weight of 3.80 ± 0.17 g and body length of 6.51 ± 1.17 cm. Four stocking density treatments were established: 50, 75, 100, and 150 individuals per cubic meter, designated as G1, G2, G3, and G4, respectively. Each treatment consisted of three replicates, resulting in a total of 12 circular tanks.
Prior to the experiment, shrimp were acclimated for two days and fed a commercial diet. The culture trial lasted 40 days and was conducted in polyethylene circular tanks (80 cm diameter, 70 cm height). Continuous aeration was provided to maintain sufficient dissolved oxygen. Shrimp were fed three times daily (08:30, 14:30, and 20:30) with a commercial formulated diet at 3–5% of total biomass. One-third of the water volume was exchanged every two days to maintain water quality.
2.2. Sample Collection and Processing
For gut microbiota analysis, shrimp were fasted for 24 h prior to sampling.
At the end of the experiment, intestines were aseptically dissected from shrimp in each treatment group.
For each biological replicate, intestines from five shrimp were pooled to obtain sufficient microbial DNA for sequencing.
All pooled samples were immediately frozen in liquid nitrogen and stored at −80 °C until DNA extraction. Samples collected at day 0 served as the control group (CK).
2.3. Growth Performance Calculations
Weight gain rate (WGR), specific growth rate (SGR), and survival rate (SR) were calculated as follows:
WGR (%) = (Final weight − Initial weight) / Initial weight × 100
SGR (% day-1) = (ln Final weight − ln Initial weight) / Culture days × 100
SR (%) = Number of surviving shrimp / Initial number × 100
2.4. DNA Extraction, Sequencing, and Bioinformatics Analysis
Total genomic DNA was extracted from intestinal samples using the E.Z.N.A.® Soil DNA Kit (Omega Bio-tek, USA). DNA quality and concentration were assessed by agarose gel electrophoresis and NanoDrop spectrophotometry. The V3–V4 region of the bacterial 16S rRNA gene was amplified using primers 338F and 806R, and sequencing was performed on an Illumina platform. Raw reads were quality-filtered, merged, and processed using QIIME2. Taxonomic assignment was conducted against the SILVA v138 database, and functional profiles were predicted using PICRUSt2.
2.5 Statistical Analysis
Growth performance and water quality data were analyzed using one-way analysis of variance (ANOVA) in SPSS software, followed by Duncan’s multiple range test.
Results are expressed as mean ± standard error (SE), and differences were considered significant at P < 0.05. For microbiome analysis, alpha-diversity indices were used to describe within-sample diversity, and beta-diversity patterns were visualized by principal coordinate analysis (PCoA) based on Bray–Curtis distances. Because no additional multivariate significance testing, such as PERMANOVA or ANOSIM, was performed in the present analysis, beta-diversity results were interpreted descriptively rather than as formally validated statistical differences. LEfSe was used as an exploratory method to identify differentially enriched taxa, and the corresponding results were interpreted cautiously rather than as independently validated biomarkers.
Results
3.1. Effects of Stocking Density on Growth Performance
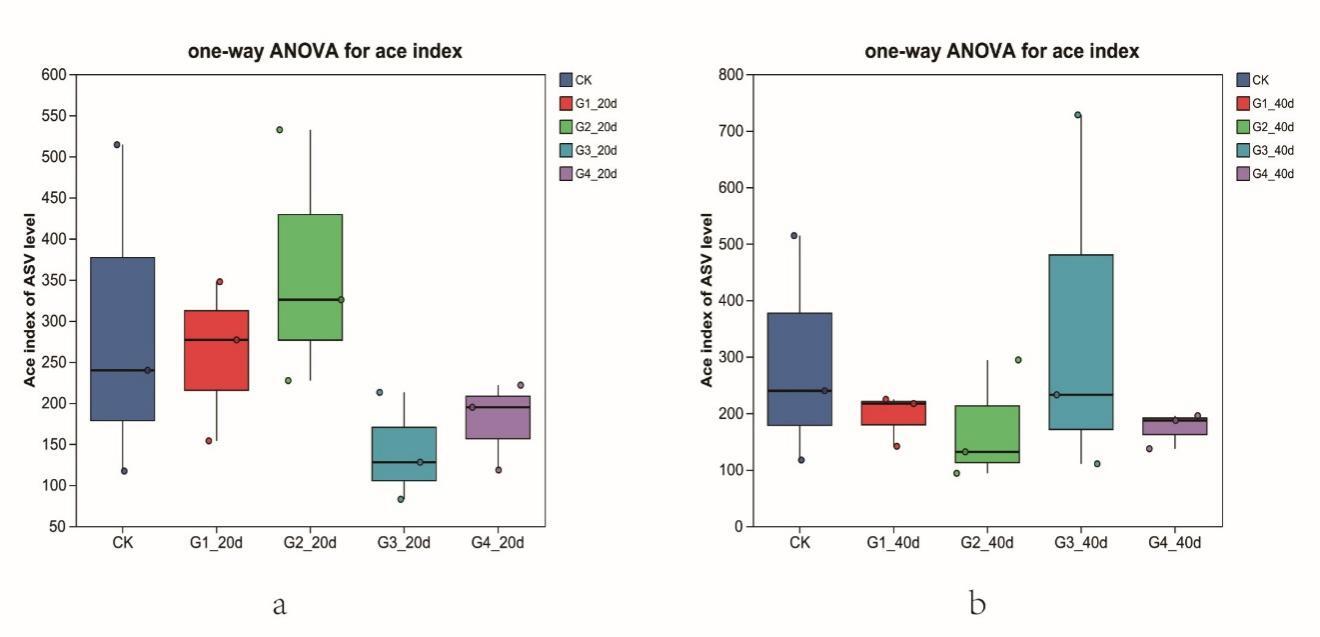
Stocking density significantly affected growth performance and survival of P. monodon (Table 1). Final body length, body weight, weight gain rate, and specific growth rate decreased progressively with increasing density. Shrimp in the low-density group (G1) exhibited significantly higher growth performance than those in the high-density groups (G3 and G4) (P < 0.05). Survival rate showed a similar trend, with the highest value observed in G1 and the lowest in G4.
3.2. Effects of Stocking Density on Water Quality
Water quality parameters remained within acceptable ranges for shrimp culture throughout the experiment (Table 2). Ammonia nitrogen and nitrite concentrations tended to increase in medium- and high-density groups during the later culture stage, while dissolved oxygen and pH remained relatively stable. Although no statistically significant differences were detected, consistent density-dependent trends were observed, particularly during the later culture stage.
3.3. Sequencing summary statistics
Sequencing summary statistics for the intestinal microbiota samples are presented in Table 3.
A total of 577,374,004 raw reads and 577,353,952 clean reads were obtained from 12 samples.
The number of raw reads per sample ranged from 40,891,838 to 61,898,742, whereas clean reads ranged from 40,890,994 to 61,896,516.
Q20 values ranged from 99.22% to 99.42%, and Q30 values ranged from 97.42% to 97.97%, indicating high sequencing quality.
The average GC content across all samples was 50.71%, ranging from 46.83% to 53.75%.
3.4. Intestinal Microbial Diversity and Community Structure
High-throughput sequencing revealed a rich and diverse gut microbiota across all treatments.
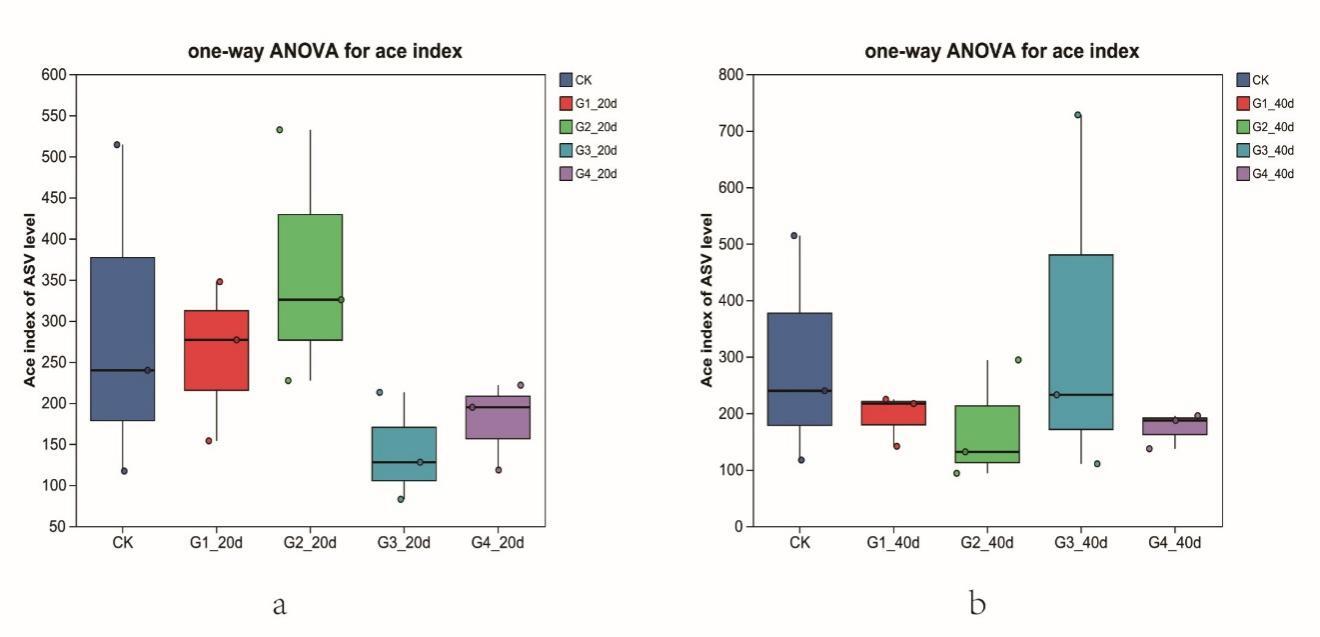
Alpha diversity indices declined with increasing stocking density (Figure 1).
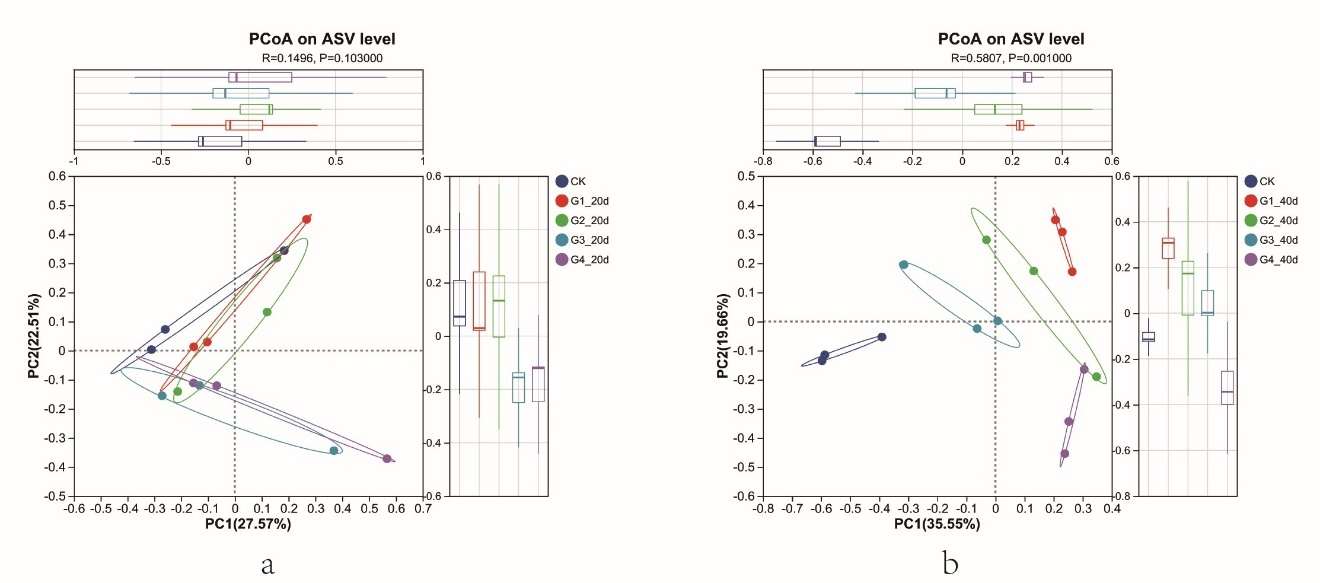
PCoA based on Bray–Curtis distances suggested differences in microbial community composition between low- and high-density treatments at day 40 (Figure 2).
_based_on_bray--curtis_distances_showing_differences_i.jpeg)
However, because no additional multivariate significance tests were performed, these beta-diversity patterns should be interpreted as descriptive compositional trends rather than definitive statistical separations.
3.5. Taxonomic Composition and Functional Prediction
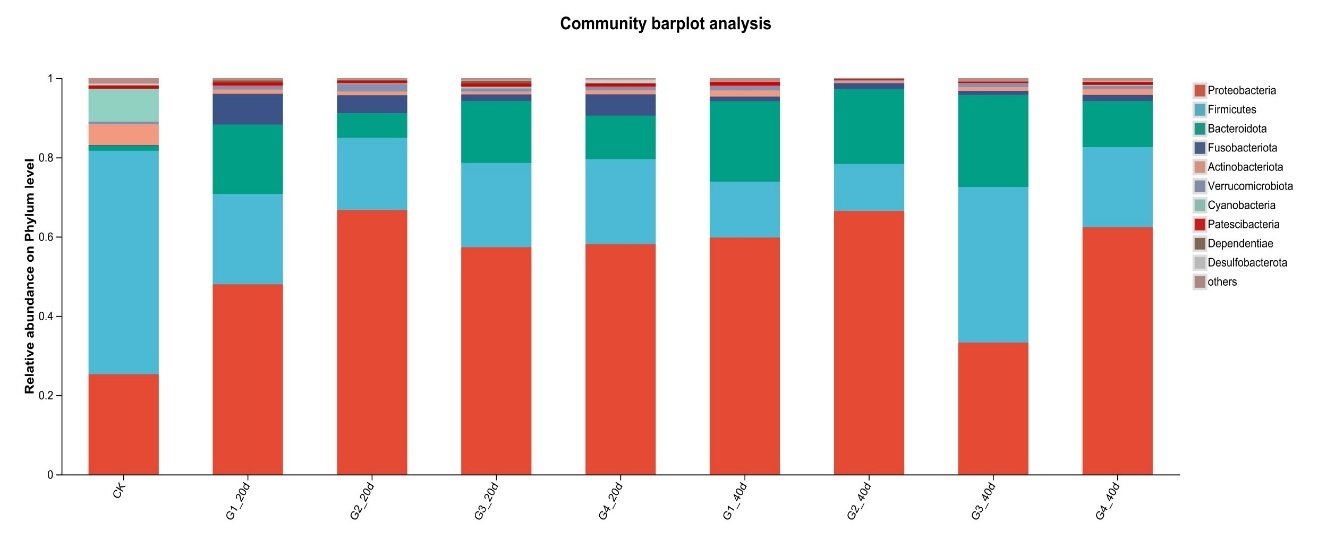
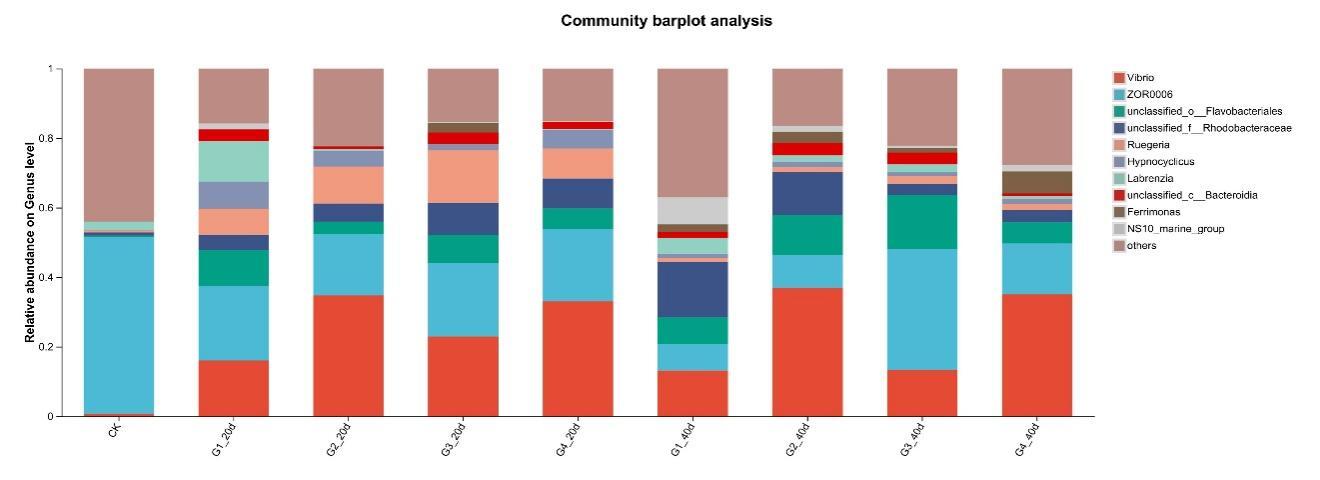
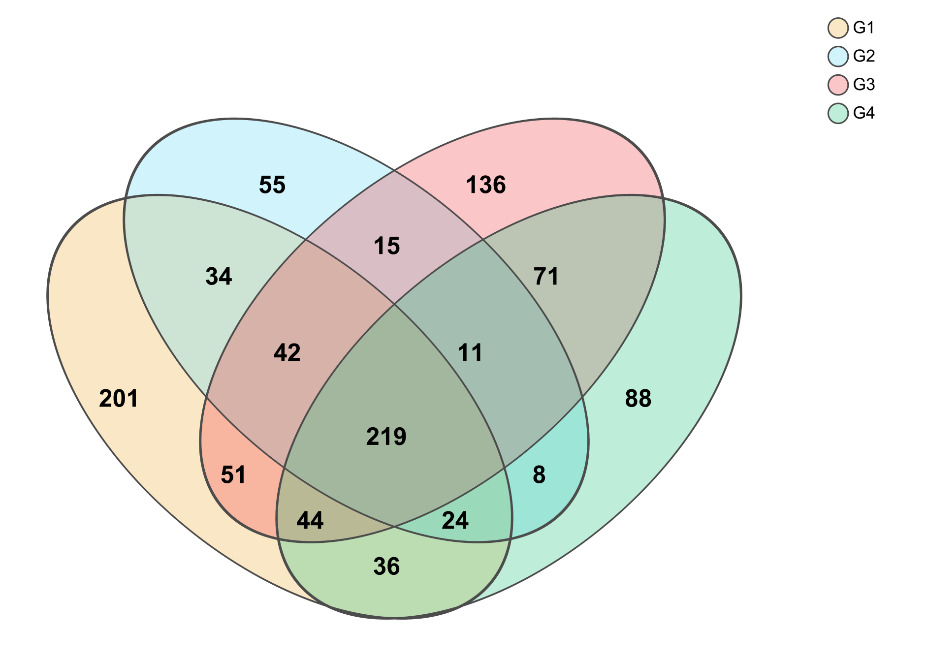
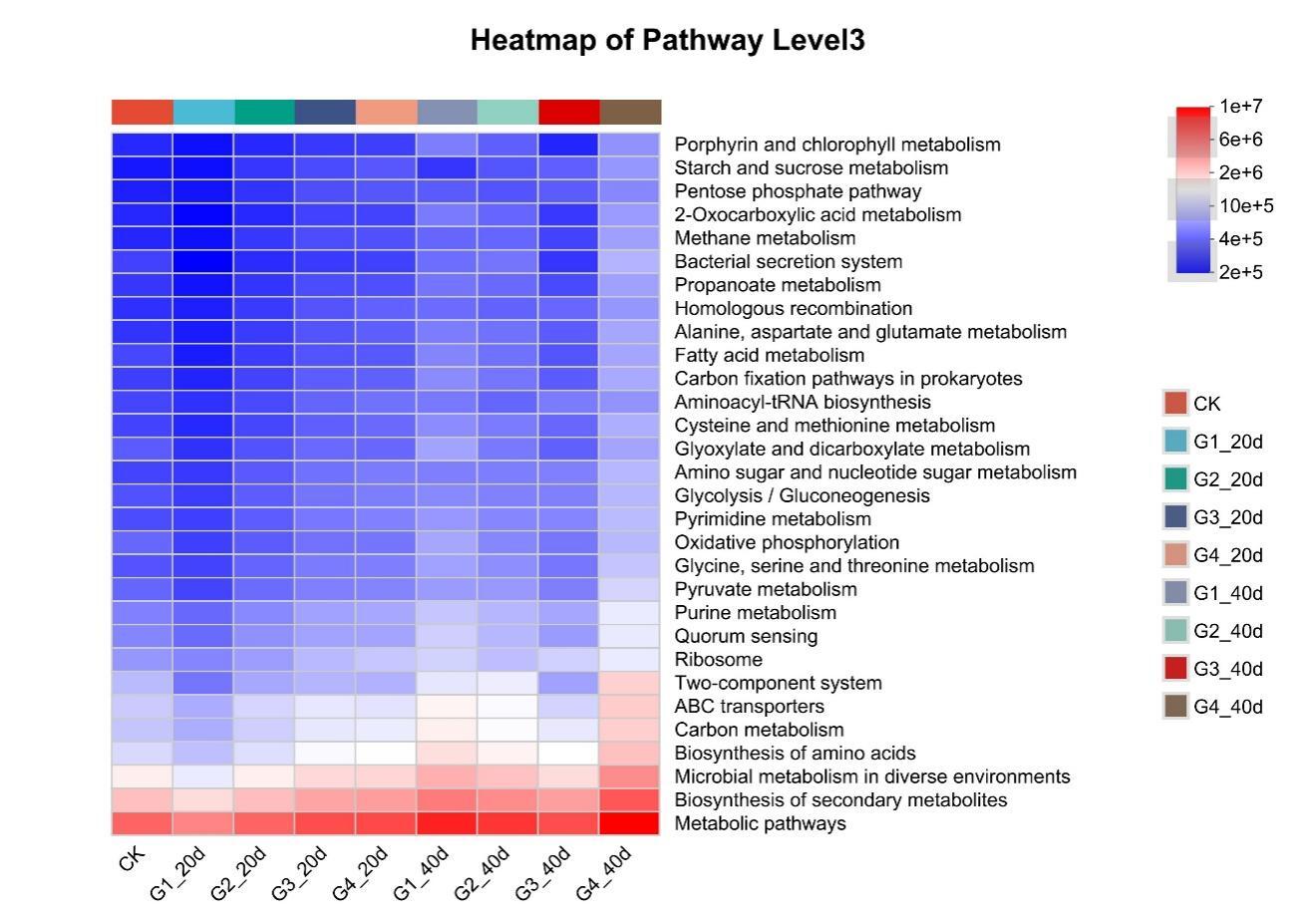
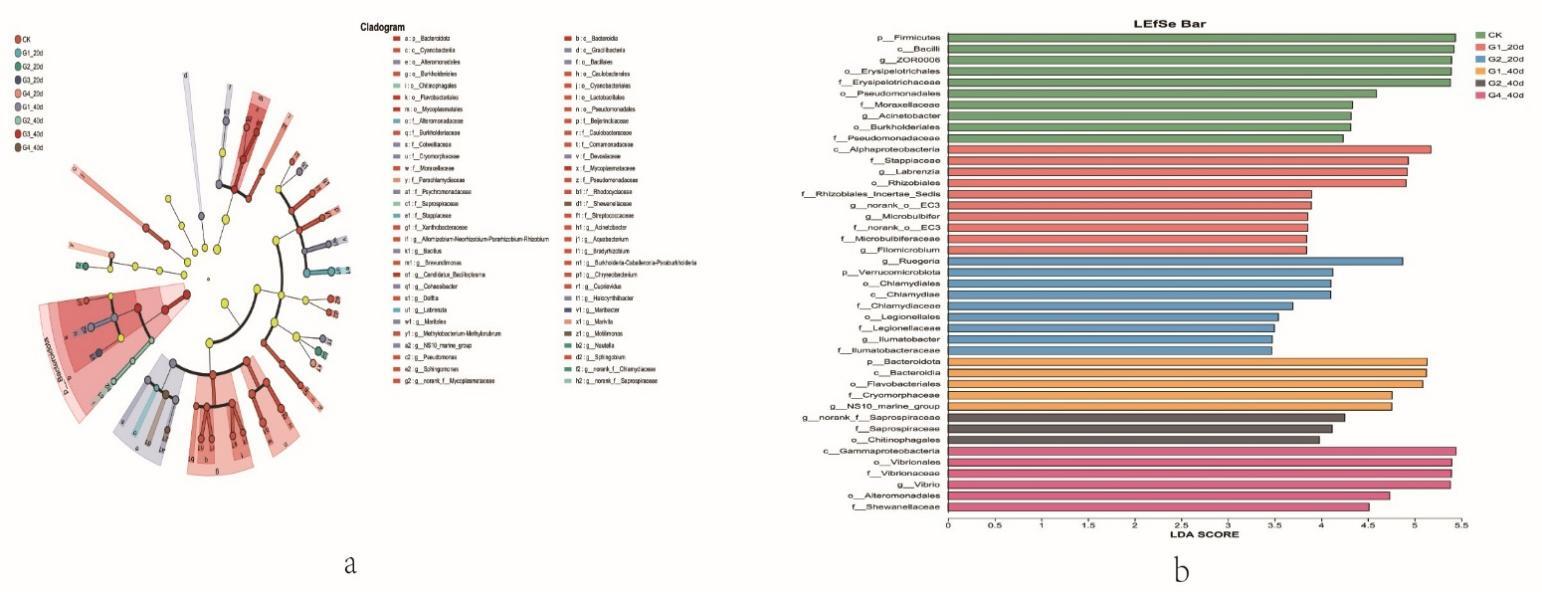
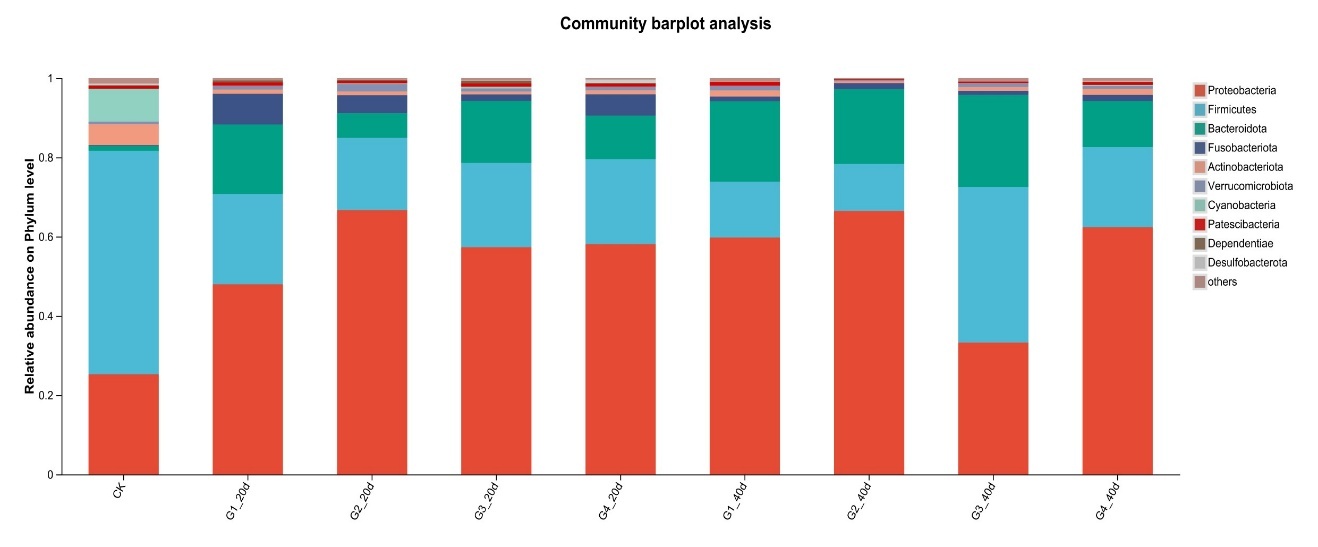
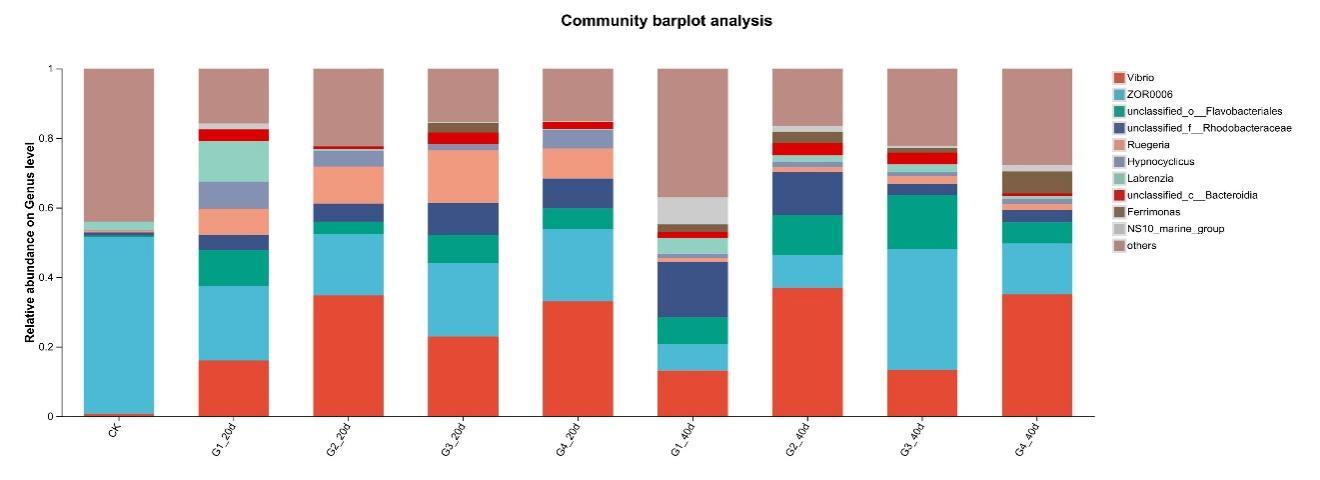
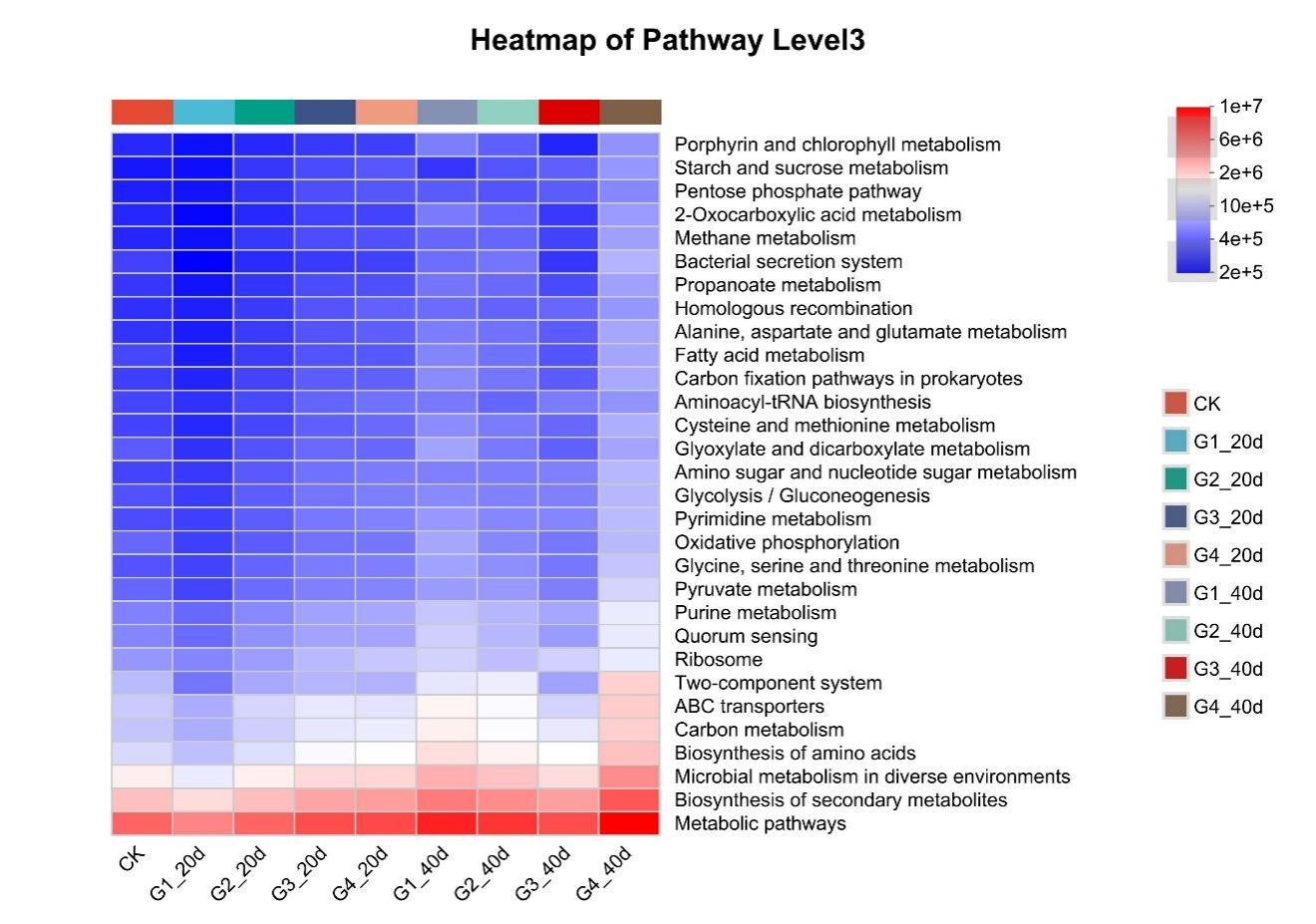
Proteobacteria dominated the gut microbiota across all treatments, followed by Firmicutes and Bacteroidota (Figure 3). High-density groups showed increased relative abundance of the potentially pathogenic genus Vibrio, whereas beneficial genera such as Bacillus and Acinetobacter were enriched in low-density groups (Figure 4). Venn analysis revealed both shared and unique ASVs among density treatments (Figure 5). Functional prediction based on PICRUSt2 indicated that high-density conditions promoted shifts toward energy-related and stress-associated metabolic pathways (Figure 6). In addition, LEfSe analysis identified distinct bacterial taxa that were significantly enriched under different stocking density treatments, highlighting density-specific microbial biomarkers associated with changes in gut community structure (Figure 7).
_among_different_s.png)
_identifying_bacterial_taxa_that_are_signi.jpeg)
Discussion
Stocking density is a fundamental management factor in intensive shrimp culture.1
In the present study, increasing stocking density significantly reduced growth performance and survival of P. monodon.4–6 Ammonia nitrogen and nitrite showed upward trends in the medium- and high-density groups during the later culture stage, suggesting that environmental loading increased as biomass and feed input accumulated over time in the culture system.10 These results indicate that the negative effects of high stocking density are better interpreted in the context of crowding and culture-system pressure in shrimp farming than by direct extrapolation from finfish physiology.11
Stocking density was also associated with variation in gut microbial diversity and community composition in the present study.
High-density treatments showed reduced alpha diversity and an altered microbial structure. These results indicate that elevated stocking density was associated with intestinal microbial shifts under indoor culture conditions.
An important limitation of the present study is that the surrounding environmental microbial community, particularly the rearing-water microbiota, was not characterized. Thus, the microbial differences observed among density treatments cannot be confidently attributed solely to stocking density, because the intestinal microbiota of shrimp is also influenced by background microbial assemblages in the culture environment.2 The intestinal microbiota of shrimp has been shown to be closely associated with microbial communities in the surrounding water and sediment.12,13 In addition, the contribution of ambient microbiota to shrimp gut microbial composition may increase during the aquaculture process. This limitation should be taken into account when interpreting treatment-driven modulation of the gut microbiome in P. monodon.
PICRUSt2 analysis suggested differences in predicted functional potential among treatments, but these results represent functional inference from 16S rRNA gene data rather than direct evidence of microbial metabolic activity.13
The relatively higher abundance of Vibrio in the high-density treatment should also be interpreted with caution. Some Vibrio species are recognized as opportunistic pathogens in shrimp aquaculture.14 However, the present study did not include pathogen isolation, species-level confirmation, virulence testing, or clinical disease diagnosis.
Therefore, the increase in Vibrio relative abundance cannot be regarded as direct evidence of pathogenic infection.
Likewise, the lower survival observed under high-density conditions should not be interpreted solely as a microbiologically driven disease outcome. Instead, reduced survival may also reflect density-related competition in P. monodon culture5 and increased environmental pressure in intensive shrimp systems.15 In contrast, Bacillus and Acinetobacter were relatively more abundant in the lower-density treatments.
The intestinal microbiota of P. monodon has been reported to vary with developmental stage and diet-related factors.7 Density-associated shifts in intestinal microbiota have also been observed in Litopenaeus vannamei.8 Taken together, these findings suggest that excessive stocking density may constrain shrimp performance through interacting effects on culture-system loading and intestinal microbial structure.
Conclusions
Stocking density is a key determinant of growth performance, water quality dynamics, and gut microbial ecology in P. monodon. Excessively high-density compromises growth, survival, and microbial stability, whereas moderate density supports balanced microbiota, stable environmental conditions, and improved production efficiency. These findings provide a scientific basis for optimizing stocking density in intensive black tiger shrimp aquaculture systems.
Acknowledgments
This research was funded by the National Key R & D Program of China (2022YFD2400104), the China Agriculture Research System of MOF and MARA (CARS-48), the Central Public-interest Scientific Institution Basal Research Fund, CAFS (No. 2023TD34 and 2025XT0703), and the Central Public Interest Scientific Institution Basal Research Fund, South China Sea Fisheries Research Institute, CAFS (No. 2024XT01). We thank the staff of the Shenzhen Experimental Base of the South China Sea Fisheries Research Institute for assistance with shrimp husbandry and sampling.
Authors’ Contribution
Conceptualization: Wenzhe Li (Equal), Jianzhi Shi (Equal). Methodology: Wenzhe Li (Equal), Jianzhi Shi (Equal). Formal Analysis: Wenzhe Li (Equal), Jianzhi Shi (Equal), Hongshan Diao (Equal), Lishi Yang (Equal). Investigation: Wenzhe Li (Equal), Jianzhi Shi (Equal), Hongshan Diao (Equal), Lishi Yang (Equal). Data curation: Wenzhe Li (Equal), Jianzhi Shi (Equal). Writing – original draft: Wenzhe Li (Equal), Jianzhi Shi (Equal). Writing – review & editing: Wenzhe Li (Equal), Jianzhi Shi (Equal). Project administration: Jiang Song (Lead). Resources: Qibin Yang (Lead). Visualization: Yundong Li (Lead). Validation: Jianhua Huang (Lead). Software: Yangyang Ding (Lead). Supervision: Falin Zhou (Lead). Funding acquisition: Falin Zhou (Lead).
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
The animal study protocol was approved by the Animal Ethics Committee of South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (protocol code nhdf2024-02; 5 February 2024). All efforts were made to minimize potential suffering during handling and sampling.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
Data will be made available on request.