

INTRODUCTION
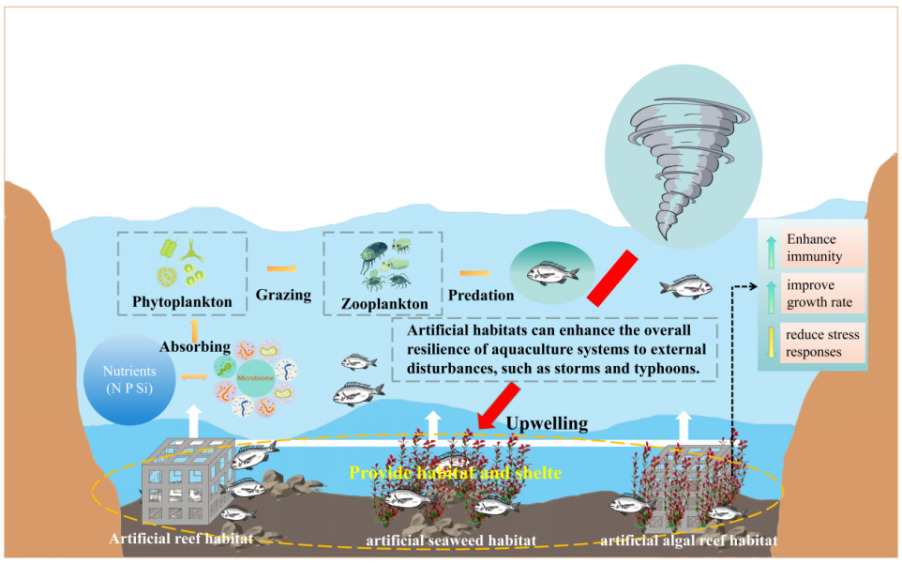
Multiple anthropogenic pressures-including coastal land reclamation, shoreline engineering, aquaculture activities, and pollutant discharge-have severely degraded fish habitats, leading to a continued deterioration of marine living environments and an accelerated decline of coastal fishery resources.1,2 In response to the need for fisheries conservation and sustainable utilization, marine ranching has increasingly become a central component of modern ecosystem-based fisheries management.3,4 Recent studies have further emphasized that marine ranching construction, shallow-water habitat assessment, and water-quality monitoring are closely associated with fishery-resource restoration and habitat-management effectiveness.5,6 As an effective approach for restoring degraded habitats and enhancing fishery resources, the deployment of artificial structures has emerged as a key strategic option in many coastal fishing nations.7,8 However, existing aquaculture systems and marine ranching habitats are often characterized by simplified structural designs and limited ecological functionality, which are insufficient to meet the diverse ecological requirements of fishes.9,10 Consequently, optimization of habitat structure has become a critical issue in stock enhancement and aquaculture research.11–13
A. schlegelii is a warm-temperate, nearshore species widely distributed along the southeastern coast of China. It is both a representative reef-associated commercial fish and an important aquaculture species in the South China Sea.14 Nevertheless, under offshore cage-culture conditions, recovery of A. schlegelii resources has remained limited. This lack of recovery is largely attributed to simplified habitat structures, high sensitivity to external environmental variability, and reduced ecosystem stability associated with cage-based farming systems.15,16
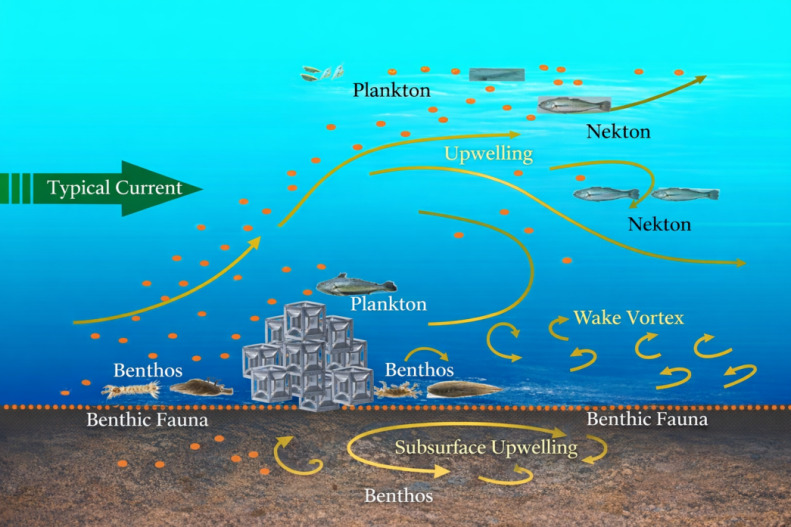
Environmental enrichment represents a suite of technical approaches aimed at improving animal welfare by optimizing habitat conditions and enhancing environmental quality.17,18 These approaches generally include physical, social, sensory, training, and nutritional enrichment.19–21 Among them, physical enrichment modifies the spatial configuration and physical attributes of rearing environments to increase habitat complexity and spatial heterogeneity, thereby influencing fish behavior, improving ecological conditions, and ultimately enhancing aquaculture performance (Fig. 1).22–24 Therefore, measurable behavioral indicators, such as spatial distribution, residence time, and swimming path length, can be used to evaluate how effectively environmental enrichment modifies habitat use and behavioral adaptation in fish.
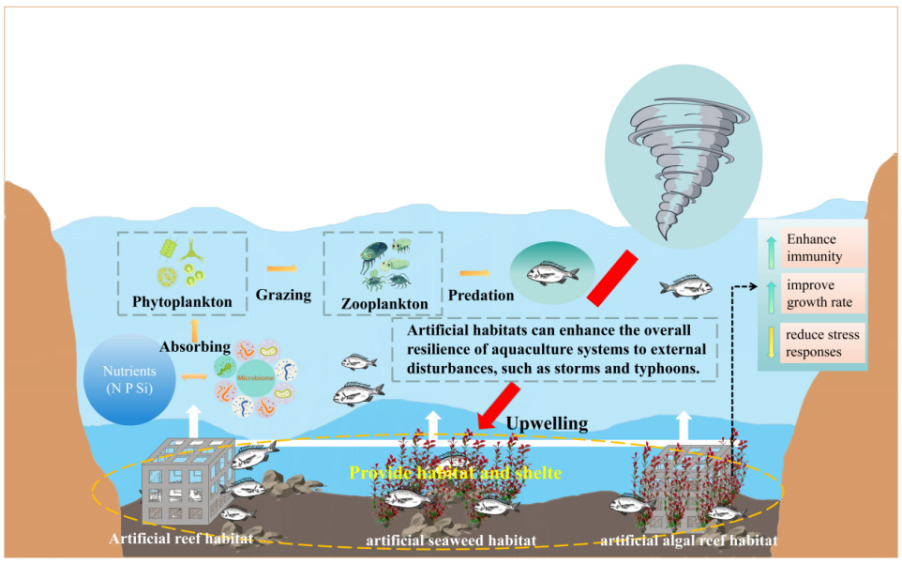
Previous studies have demonstrated that different forms of structural enrichment can significantly alter fish behavior. For example, Jiang et al.25 examined behavioral preferences of juvenile redfin perch toward artificial reef models with varying aperture shapes and sizes, while Sass et al.26 investigated the effects of coarse woody habitat additions on population dynamics, feeding behavior, and habitat use of Lepomis macrochirus and Micropterus salmoides bass. However, most existing research has focused on artificial reef with single structural forms, whereas studies addressing composite reef structures that more closely resemble natural rocky reef, as well as habitats simulating seaweed, remain limited.27,28 Therefore, the present study employs controlled laboratory-based behavioral observation experiments to examine the selective preferences of A. schlegelii for artificial seaweed reef, artificial seaweed, and artificial reef across four diel periods (0 - 6 h, night; 6 - 12 h, morning; 12 - 18 h, afternoon; 18 - 24 h, evening). By quantitatively characterizing adaptive behavioral responses of A. schlegelii under different structured habitats, this study aims to provide technical support for habitat enrichment in A. schlegelii aquaculture and for the conservation and management of fishery resources (Fig. 2).18

MATERIALS AND METHODS
MATERIALS
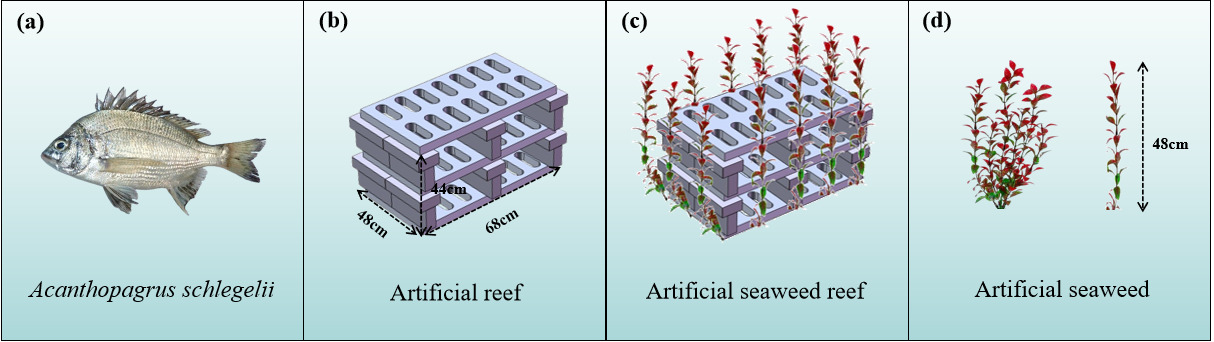
A. schlegelii used in the experiment were obtained from a commercial aquaculture facility in the coastal waters of Zhoushan, China. The experimental individuals had a mean total length of 30.0 ± 0.5 cm, body height of 5.01 ± 0.5 cm, and body mass of 486.0 ± 50.0 g. The artificial reef used in the experiment consisted of a hollow concrete structure. The artificial seaweed (Fig. 3) exhibited a morphology comparable to that of natural macroalgae, forming a clustered, canopy-like structure, and was manufactured from non-toxic, odorless polyethylene materials.
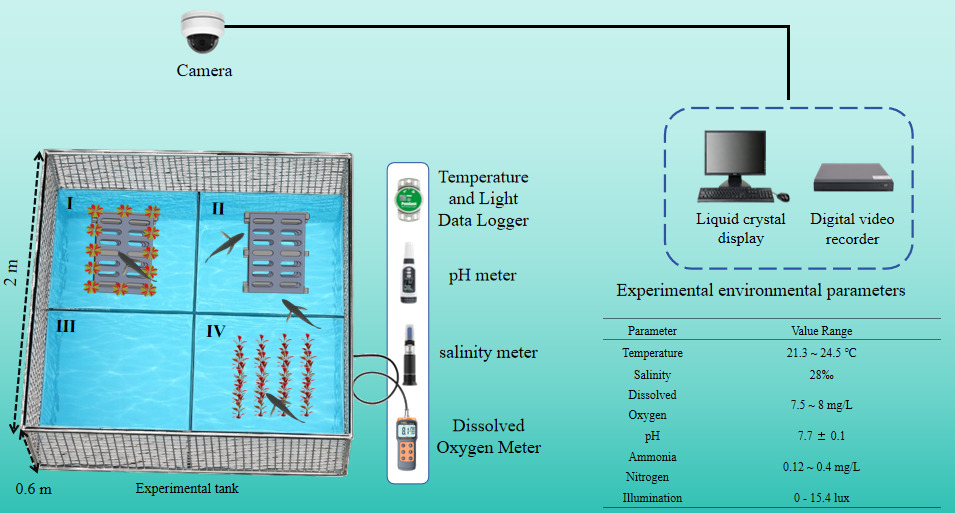
Prior to the experiment, the artificial seaweeds were soaked in seawater for two weeks and repeatedly rinsed to remove surface debris and potential contaminants. Before formal observations, A. schlegelii were transferred to the experimental tank and acclimated for 24 h without feeding to minimize stress-related effects on behavior. Water quality parameters during the experiment are shown in (Fig. 4). Experimental equipment included a Hikvision high-definition infrared video monitoring system, XiMa pH828 pH meter (measurement range: 0.00 - 14.00), Delixi salinity meter (measurement range: 0 - 100‰), dissolved oxygen meter, HOBO wireless Bluetooth underwater temperature and light logger, and a computer.

HABITAT MODELS AND TANK ZONING DESIGN
Four habitat treatments were established to examine habitat use by A. schlegelii under different structural conditions: Artificial reef, artificial seaweed, artificial seaweed reef, and barren water control. In this study, “structured habitats” refer to treatments containing artificial reef, artificial seaweed, or their combined structures, whereas “structural complexity” describes the degree of physical heterogeneity created by these artificial structures. The artificial reef consisted of stacked grey bricks forming a rectangular structure (68 × 48 × 44 cm; length × width × height) with internal openings measuring 27 × 16 cm. Artificial seaweed units (48 cm in height) were arranged over a 50 × 50 cm area with a spacing of 15 cm between units. The artificial seaweed reef comprised an artificial reef surrounded by artificial seaweed arranged at the same spacing. Schematic representations of the habitat structures are provided in (Fig. 3). Experiments were conducted in a rectangular tank (200 × 200 × 60 cm) with a water depth of 50 cm. The tank was cleaned and disinfected prior to experimentation, and the bottom was marked at 1 m intervals using green reference points to delineate four equal-area zones (1 m² each). Zones I - IV corresponded to the artificial seaweed reef, artificial reef, barren water control, and artificial seaweed treatments, respectively (Fig. 4). As fish positions were assigned according to horizontal zone boundaries, this experiment primarily assessed horizontal habitat use rather than three-dimensional vertical distribution.

EXPERIMENTAL DESIGN
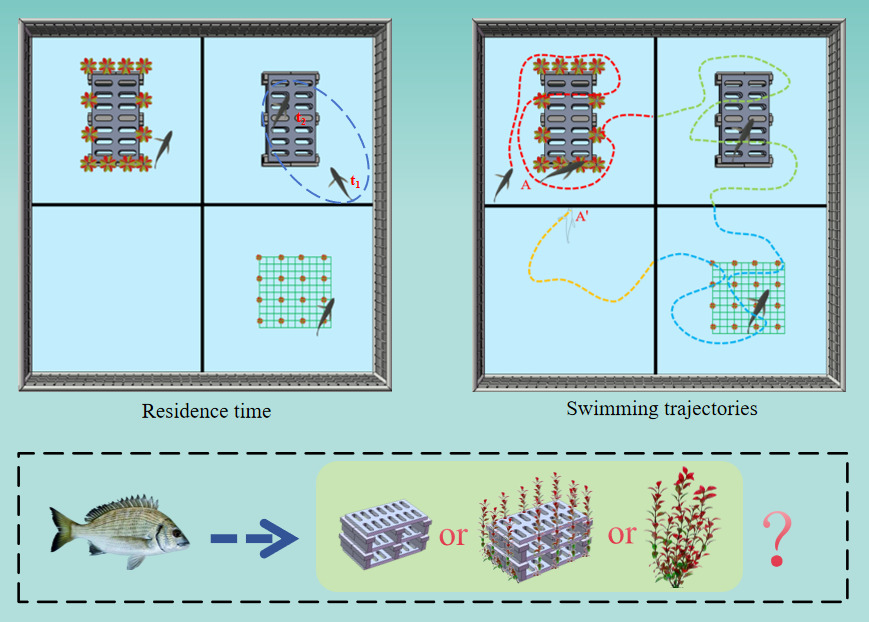
Habitat selection experiment: To examine spatial distribution patterns of A. schlegelii under contrasting habitat conditions, the experimental tank was divided into four equal-area habitat zones using predefined spatial boundaries (2.0 × 2.0 × 0.6 m, length × width × height). Each arena was subdivided into four equal-area blocks (Zones I - IV), which were assigned to barren water control, artificial seaweed, artificial reef, and artificial seaweed reef treatments, respectively. Shading curtains were deployed to minimize external light interference, and the experimental environment was kept quiet to reduce disturbance. Prior to data collection, four individuals were introduced into each enclosure and were allowed to acclimate for 6 h. Following acclimation, continuous behavioral observation was conducted. Video was recorded continuously for 3 consecutive days (00:00 - 24:00), and one frame was extracted every 5 min to document the spatial distribution. Habitat occupancy was determined according to predefined zone boundaries; when an individual was located on a boundary, assignment was based on head position. Cumulative counts of fish occurrences per zone were used to compare distributional differences and to evaluate habitat preference in a behavioral observation experiment. Within the constructed experimental arenas, the entire experimental process was continuously video-recorded for subsequent behavioral analyses:
(1) Swimming path analysis: Using Tracker v6.2.0, samples were drawn at 2 h intervals over a 24 h cycle. For each sampling time, a continuous 3 min segment beginning at the interval start was analyzed. Habitat affiliation was assigned based on the position of the fish head relative to zone boundaries, and swimming path length was measured from the time an individual entered a zone until it exited. Trajectories of all individuals entering a zone were superimposed, and mean swimming path lengths were computed for each fish across time periods and habitat types.
(2) Residence-time analysis: Residence times were quantified by replaying 2 min video segments (recorded every 2 h) at reduced speed in VLC Media Player and recording the duration that individuals remained within each zone. Spatial distribution and behavioral metrics were then compared among habitat types to assess habitat-specific behavioral adaptation (Fig. 5).

DATA ANALYSIS
The number of fish occurrences, swimming path length, and residence time within each habitat zone were used as primary quantitative indicators of behavioral responses. Behavioral data were systematically recorded and compared across different time periods to evaluate habitat preference and its temporal dynamics. All statistical analyses were performed using Excel and IBM SPSS Statistics 27. Results are presented as mean ± standard deviation (Mean ± SD). Differences among groups were assessed using one-way analysis of variance (ANOVA) and independent-sample t-tests, with statistical significance set at 0.05. The calculation methods for mean occurrence frequency, swimming path length, and residence time were as follows:
(1) Mean occurrence frequency: The total number of occurrences of A. schlegelii in a given habitat was recorded separately for four equal observation periods (each 6 h). The mean occurrence frequency for that habitat was reported as the arithmetic average of the four period totals.
(2) Mean swimming path length: Swimming distance was measured in 5 min observation windows. For each fish, nine 5 min measurements (three replicate sampling times across three sampling periods) were obtained, and their arithmetic mean was taken as the fish’s mean swimming path length. Habitat-level mean path lengths were then derived from these per-fish averages.
(3) Mean residence time: Residence time was defined as the duration of a single stay within the target habitat. For each fish, three repeated observations of single-stay duration were recorded, and the mean of these three values was used as the fish’s mean residence time; habitat means were computed from individual means.
RESULTS
HABITAT-SPECIFIC DISTRIBUTION
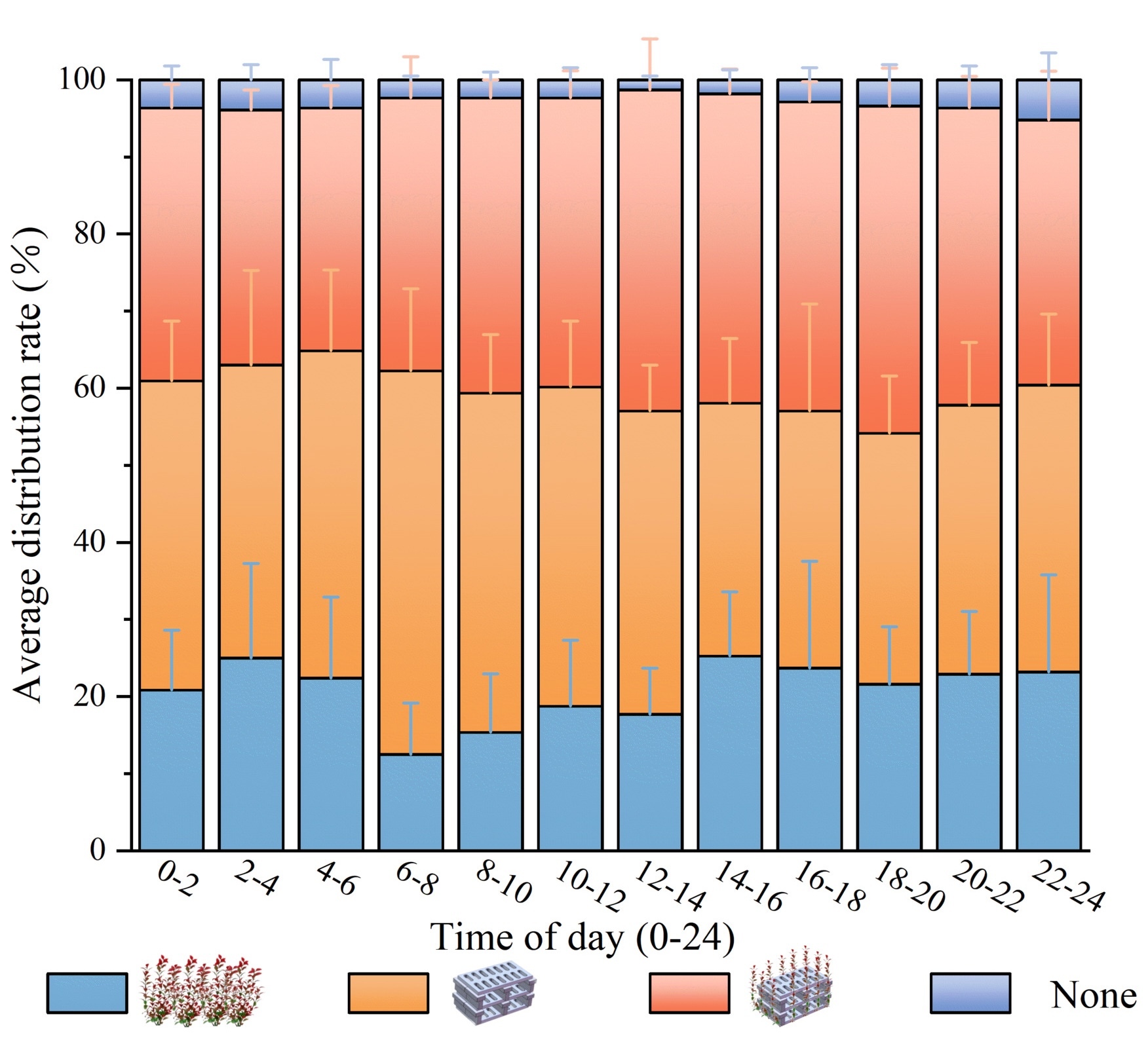
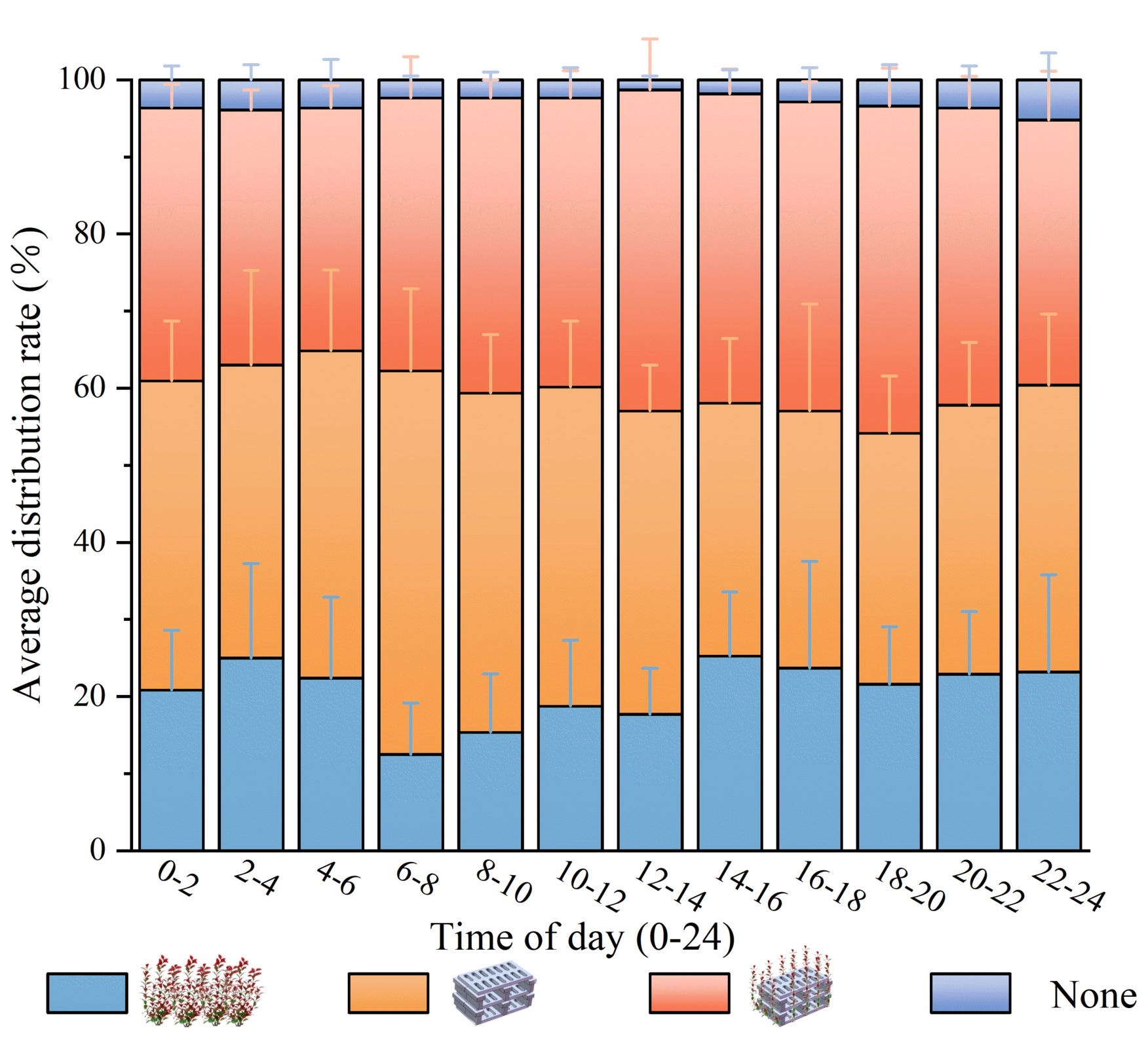
Fig 6 shows the distribution of A. schlegelii across the four habitat types. Statistical analysis indicated that habitat type had a significant effect on occurrence frequency (p < 0.05). Overall, mean occurrence frequencies were highest in the artificial reef (37.27 ± 10.31 occurrences) and the artificial seaweed reef (35.94 ± 5.35 occurrences); these two treatments did not differ significantly from one another but were both significantly greater than the artificial seaweed (19.94 ± 8.75 occurrences) and the barren water control (2.92 ± 1.94 occurrences) (p < 0.05). Temporal patterns differed among habitats: the artificial reef exhibited a morning aggregation peak (06:00 - 08:00; 47.75 ± 8.50 occurrences) that was significantly greater than the artificial seaweed reef during the same interval, whereas the artificial seaweed reef reached its maximum attraction at midday (12:00 - 14:00; 40.00 ± 10.80 occurrences). Occurrence frequencies in the artificial seaweed remained comparatively low across all time periods (14 - 24 occurrences) and were significantly lower than those in both artificial reef types. The barren water control consistently showed the lowest occurrence frequency (1 - 5 occurrences) and differed significantly from all structured habitats. Artificial reef structures produced a stronger aggregation effect on A. schlegelii than artificial seaweed, and the attraction exerted by artificial reef and artificial seaweed reef exhibited complementary diel timing.
RESIDENCE TIME DIFFERENCES AMONG HABITATS
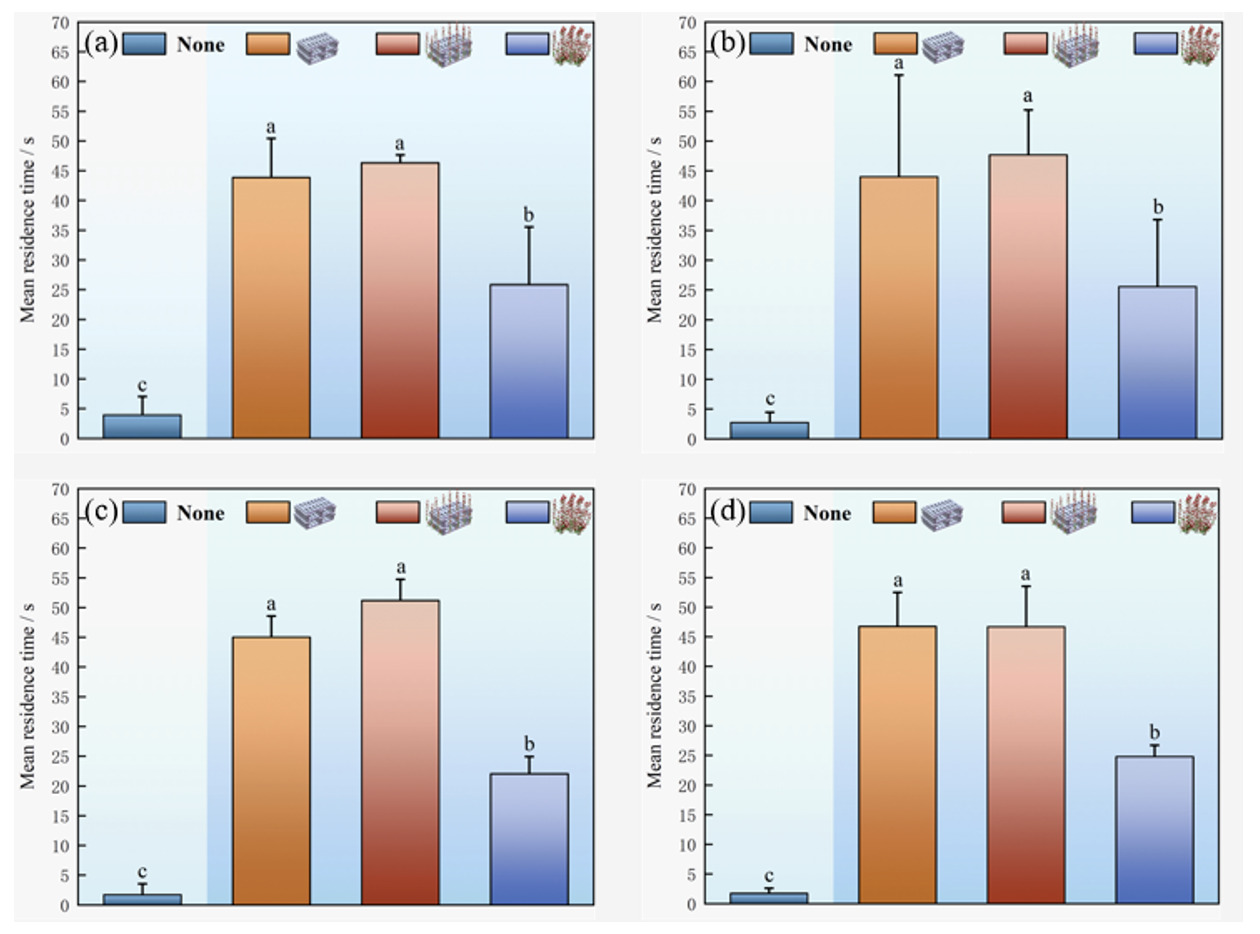
Analysis of mean residence time aggregated over three consecutive days indicates that habitat type had a significant influence on residence behaviour (p < 0.05). Structured habitats elicited substantially longer residence times than the barren water control. Among structured treatments, the artificial reef and the artificial seaweed reef produced the strongest residency effects, with the latter occupying an intermediate position. Residence times in the open control were generally short (mostly < 10 s) and showed limited variability (Fig. 7).
Between structured habitats, the mean residence time was highest in the artificial reef, followed by the artificial seaweed reef and then the artificial seaweed. In the artificial reef zone, mean residence time reached 50.25 ± 15.38 s, which was significantly greater than residence in the artificial seaweed reef and artificial seaweed (p < 0.05), and reached a nocturnal peak at 22:00 (77.25 s). Residence time in the artificial seaweed reef was likewise significantly longer than in the artificial seaweed and barren water, indicating that the composite structure of the artificial seaweed reef enhanced retention and behavioral stability.
Habitat identity exerted a stronger influence on residence behaviour than temporal factors. Habitat attraction decreased from artificial reef to artificial seaweed reef, then to artificial seaweed, and was lowest in barren water. Overall, the structural complexity and shelter availability are key drivers of residence-time allocation, and more structurally complex and functionally complete artificial habitats promote the formation of stable residency patterns in A. schlegelii.

SWIMMING PATH LENGTH AMONG HABITATS
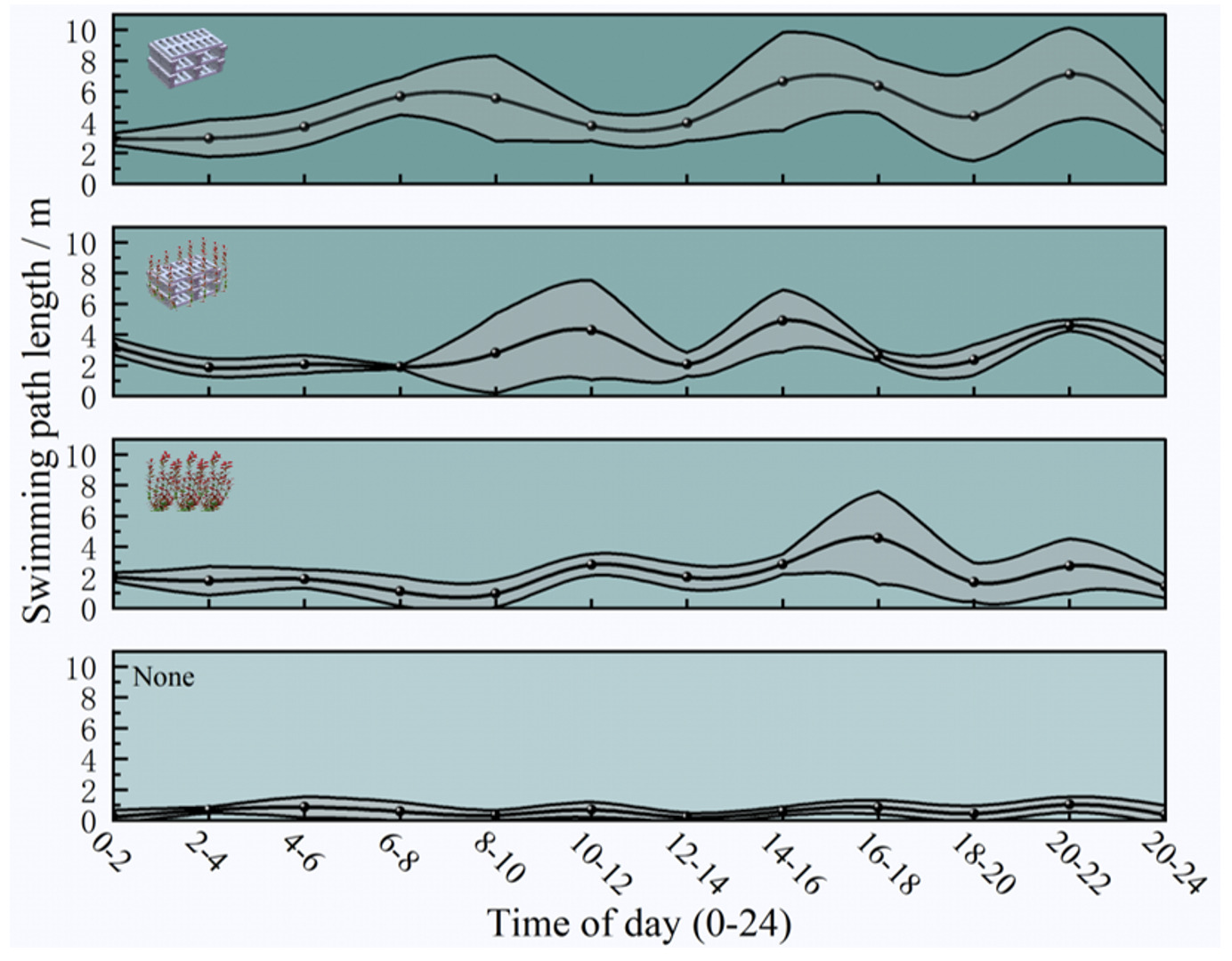
As shown in (Fig. 8), the mean swimming path length of A. schlegelii varied among habitats across time periods. The artificial reef habitat exhibited the longest mean path lengths, with values ranging from 2.64 to 10.05 m. The artificial seaweed reef was intermediate (1.40 - 8.73 m), artificial seaweed occupied an intermediate-low range (0 - 7.40 m), and the barren water control showed the shortest paths (predominantly < 1.0 m; range 0 - 3.70 m). Path lengths in the barren water treatment were significantly lower than those in the three structured habitats across time periods (p < 0.05).

Mean swimming path length varied among habitat types and exhibited clear diel patterns (Fig. 8). Across all time intervals, the longest paths were consistently recorded in the artificial reef habitat, followed by the artificial seaweed reef and artificial seaweed habitats, whereas values in the barren water control remained lowest throughout the day.
During the nocturnal period (00:00 - 06:00), mean path lengths were moderate in structured habitats, ranging from approximately 2.6 to 4.9 m in the artificial reef, 1.4 to 3.7 m in the artificial seaweed reef, and 1.5 to 2.6 m in artificial seaweed, while movement in barren water remained limited. Activity increased during the morning period (06:00 - 12:00), particularly in the artificial reef, where path lengths reached their first daily peak, and in the artificial seaweed reef, which showed a marked increase toward midday.
The highest levels of swimming activity occurred during the afternoon (12:00 -18:00), with the artificial reef exhibiting the greatest path lengths (approximately 6 - 10 m), and the artificial seaweed reef and artificial seaweed showing intermediate values. During the evening period (18:00 - 24:00), swimming activity declined across all habitats, although path lengths in the artificial reef remained relatively elevated compared with other habitats. Barren water consistently supported the shortest swimming paths at all time intervals (Fig. 9).

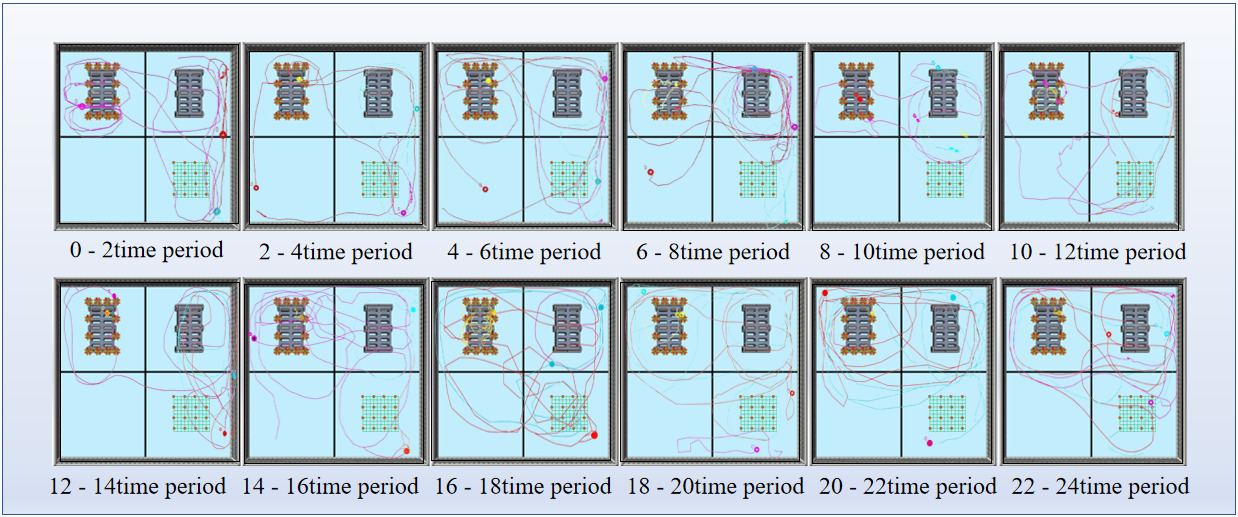
Comparison of trajectory plots across time periods reveals pronounced differences in spatial use (Fig. 10): The artificial reef substantially promoted nocturnal residency, whereas the artificial seaweed reef favored expansion of activity range and spatial utilization around midday. The two artificial reef types exhibit temporally complementary functional roles.

DISCUSSION
Habitat selection and behavioral adaptation represent key ecological strategies by which fishes optimize energy allocation, reduce predation risk, and enhance survival success in heterogeneous environments.29,30 Structured habitats increase spatial heterogeneity and refuge availability, thereby providing diversified spaces for resting, sheltering, and predator avoidance (Fig. 11). These features can reduce environmental stress, promote the expression of natural behaviors, and improve energetic efficiency.31,32 In recent years, fish welfare has become an increasingly important issue in aquaculture and fisheries resource management.33 Environmental enrichment, particularly physical enrichment aimed at increasing habitat structural complexity, has been recognized as an effective approach to improving fish welfare.34 Previous studies have shown that structurally complex artificial habitats can reduce predation risk, decrease non-functional swimming and stress-related behaviors, and enhance residency stability and behavioral diversity - responses widely regarded as indicators of improved fish welfare.35,36

The present results demonstrate that habitat structural type significantly influences the spatial distribution, locomotor behaviour and residency characteristics of A. schlegelii (p < 0.05). Mean occurrence frequency was highest in the artificial reef (37.27 ± 10.31) and artificial seaweed reef (35.94 ± 5.35) treatments, and substantially lower in the artificial seaweed treatment (19.94 ± 8.75) and the barren-water control (2.92 ± 1.94). Temporal patterns further distinguished structured habitats: artificial reef exhibited a pronounced morning aggregation peak (06:00 - 08:00; 47.75 ± 8.50), whereas artificial seaweed reef reached maximum attraction at midday (12:00 - 14:00; 40.00 ± 10.80). Residence and locomotor metrics paralleled these distributional patterns - mean nocturnal residence in the artificial reef reached 50.25 ± 15.38 s (peak 77.25 s), and mean swimming-path lengths were greatest in the artificial reef (2.64 - 10.05 m), remained substantial in the artificial seaweed reef (1.40 - 8.73 m), and were lowest in barren water (0 - 3.70 m). Overall, occurrence, residence, and movement data indicate significantly stronger preferences for artificial reef and artificial seaweed reef than for artificial seaweed, consistent with the ecological function of artificial structures in providing shelter, foraging opportunities, and predator-avoidance space.38,39 Previous work suggests that reefs with cavities, crevices and multi-dimensional sheltering promote aggregation and stable residency. These composite seaweed-reef configurations more closely resemble natural reef-seaweed mosaics and thus enhance attraction efficiency.40,41 These findings accord with Helder et al.42 who reported strong effects of structural complexity on fish spatial distribution, and with Lee who observed concentration of A. schlegelii within reef interiors compared with open controls.
From a behavioral-adaptation perspective, residence-time analyses further indicated that A. schlegelii exhibited significantly longer residency in structured habitats than in barren water (p < 0.05). Mean residence time followed a general gradient, with the artificial seaweed reef supporting the longest overall residency, followed by the artificial reef and artificial seaweed. Notably, although the artificial reef did not have the highest overall residence time, it showed a pronounced nocturnal residency, averaging 50.25 ± 15.38 s and peaking at 77.25 s at night. In contrast, residence in artificial seaweed was shorter and more variable, while residency in barren water remained minimal. This pattern may be associated with differences in structural complexity and ecological functionality among habitats. By mimicking a natural reef-seaweed composite habitat, artificial seaweed reef provides more continuous shelter and activity space, thereby promoting prolonged residency and more stable spatial use.43 Similar studies have shown that residence time of sparid fishes increases several-fold following artificial reef deployment, while remaining extremely short under non-reef conditions.44,45 In addition, fish may exploit reef and seaweed components in different ways. For example, rigid reef structures primarily function as resting and sheltering sites, whereas seaweed canopies facilitate foraging and expansion of activity space.46
Mean swimming path length differed significantly among habitat types (p < 0.05), with the highest values consistently observed in the artificial reef, followed by the artificial seaweed reef, intermediate values in artificial seaweed, and the lowest values in barren water. Quantitatively, swimming path lengths in the artificial reef ranged from 2.64 to 10.05 m across time periods, compared with 1.40 - 8.73 m in the artificial seaweed reef, 0 - 7.40 m in artificial seaweed, and predominantly less than 1.0 m (maximum 3.70 m) in barren water. This pattern indicates that structured habitats can effectively regulate activity intensity and spatial use in A. schlegelii.47,48 Sparid fishes generally prefer rocky reefs, mixed substrates, and structurally complex habitats containing crevices and cavities, which are used for both foraging and predator avoidance.49,50 The present study further revealed pronounced diel variation in swimming path length, with higher values during daytime and lower values at night. Mean path lengths increased during daylight hours, reached afternoon maxima (up to 10.12 m in the artificial reef between 14:00 and 16:00), and declined during the night across all habitats. This pattern is consistent with behavioral rhythms jointly regulated by endogenous biological clocks and light conditions.51,52 Enhanced daytime illumination facilitates visually mediated foraging and spatial exploration, whereas reduced nighttime visibility promotes activity suppression and aggregation within shelter structures.53,54 Because barren water habitats lack refuge, A. schlegelii maintained low activity levels throughout most of the day. In contrast, the stronger three-dimensional structure and sheltering capacity of artificial reefs supported the longest swimming paths during periods of elevated daytime activity and sustained relatively stable spatial use at night. Activity levels in artificial seaweed reefs were slightly lower, potentially reflecting partial obstruction of reef openings by artificial seaweed.55,56
In the present study, distribution frequency, swimming path length, and residence time of A. schlegelii were all significantly higher in artificial reef and artificial seaweed reef habitats than in artificial seaweed and barren water. These quantitative patterns suggest that combining rigid reef structure with surrounding seaweed elements provides both shelter and daytime foraging, space-use opportunities, and a structural and functional mosaic that neither component alone fully delivers. These behavioral responses reflect active selection for shelter and spatial resources, highlighting the positive role of structured habitats in regulating behavioral states and spatial utilization. The present findings provide behavioral-level evidence supporting the ecological value of artificial reefs and artificial seaweed reefs in optimizing habitat conditions, enhancing fish welfare, and promoting habitat restoration for A. schlegelii .57–59
Despite demonstrating the advantages of artificial seaweed reef, artificial reef, and artificial seaweed in improving habitat conditions, reducing stress-related behavior, and enhancing welfare, knowledge remains limited regarding how A. schlegelii respond to artificial structures differing in configuration, color, and material. Further investigation into the materials and construction of artificial structures is essential for understanding behavioral adaptation and survival enhancement of adult A. schlegelii in simulated ecological environments.60,61 Moreover, differences exist between controlled laboratory settings and natural marine environments. In particular, tank confinement may restrict swimming range and alter movement trajectories, factors that should be considered when extrapolating laboratory-derived behavioral patterns to open-water conditions. Future studies should integrate scuba-based observations and underwater video monitoring to assess fish aggregation around deployed artificial reefs and attached artificial seaweed in situ, determine effective influence radii of reef and seaweed habitats, and combine field observations with laboratory attraction experiments.62 Such integrative approaches will provide guidance on reef material selection, structural design, and the construction of artificial seaweed habitats.
ACKNOWLEDGMENTS
This research project was funded by the National Key Research and Development Program of China (2025YFE0113200), National Science Foundation of China (32503264).
AUTHORS’ CONTRIBUTION – CREDIT TAXONOMY
Conceptualization: Xunmeng Li (Lead). Data curation: Hailong Wang (Equal), Kai Zong (Equal). Methodology: Jinhuai Ni (Lead). Formal Analysis: Kai Zong (Lead). Investigation: Hailong Wang (Lead). Writing - original draft: Hailong Wang (Lead). Writing - review & editing: Xunmeng Li (Lead). Funding acquisition: Xunmeng Li (Lead). Supervision: Lingling Zhang (Lead). Software: Jianpeng Lu (Lead). Validation: Zhaocong Lansun (Lead). Project administration: Fukun Gui (Lead).
COMPETING OF INTEREST - COPE
No competing interests were disclosed.
ETHICAL CONDUCT APPROVAL - IACUC
Ethical approval is not required for this research.
INFORMED CONSENT STATEMENT
All authors and institutions have confirmed this manuscript for publication.
DATA AVAILABILITY STATEMENT
The data that has been used is confidential.