
Introduction
Coral reef ecosystems are currently facing an unprecedented global decline, driven by the synergistic effects of anthropogenic climate change, overfishing, and localized nutrient pollution.1 As natural recovery rates are increasingly outpaced by the frequency of bleaching events and habitat degradation, active restoration strategies have become a cornerstone of marine conservation.2 The “coral gardening” concept, which involves the nursery cultivation of fragments before their transplantation back to degraded reefs, has proven effective in bypassing the high-mortality early life stages of corals.3 Recently, land-based Recirculating Aquaculture Systems (RAS) have emerged as an essential infrastructure for ex-situ coral propagation, offering precise control over environmental parameters and providing a biosecure environment for the mass production of coral biomass.4,5
A significant technical advancement in coral aquaculture is the application of micro-fragmentation. This technique involves cutting coral colonies into minute fragments (typically 1–3 cm² or linear extension), which triggers a vigorous physiological “healing response” and stimulates rapid tissue expansion to overgrow the exposed substrate.6 While micro-fragmentation maximizes the output from limited donor material, maintaining the high metabolic demand of these rapidly growing fragments requires an optimized nutritional balance between autotrophy (photosynthesis) and heterotrophy (external feeding). Scleractinian corals, particularly those in the genus Acropora, are highly plastic in their energy acquisition, capable of supplementing their energy budget through the ingestion of particulate organic matter, zooplankton, and the absorption of dissolved organic matter.7 Heterotrophic feeding has been shown to significantly enhance chlorophyll-a concentrations, skeletal density, and the accumulation of protein and lipid reserves, which are critical for the long-term resilience of the fragments8,9
However, a critical unresolved question remains regarding the trade-off between intensive heterotrophy and water quality stability in closed-system aquaculture. Despite the clear physiological benefits of heterotrophy, intensive feeding in closed RAS environments presents substantial management challenges. Excessive organic loading can lead to a rapid accumulation of dissolved inorganic nutrients, particularly phosphate (PO4), which is known to inhibit aragonite precipitation and skeletal calcification in many Acropora species.10 Furthermore, nutrient enrichment frequently facilitates the proliferation of opportunistic filamentous algae and cyanobacteria. These competitors can rapidly outgrow coral micro-fragments, leading to competitive exclusion, tissue smothering, and localized hypoxia at the coral-algal interface.11,12 While the role of trace elements—such as Strontium (Sr), Iodine (I), and Iron (Fe)—as biochemical catalysts for calcification is documented,13 it remains unknown whether systematic trace element supplementation can physiologically buffer corals against the stressors of high-dose organic loading.
This nutritional challenge is further complicated by “morphological determinism.” The efficacy of nutrient uptake and waste removal is fundamentally modulated by the colony’s architecture, which creates a specific hydrodynamic environment. Tabular corals, such as Acropora hyacinthus, possess a horizontal, flat architecture that maximizes light interception for photosynthesis but acts as a “catch-basin” for organic debris and algal settlement.14 In contrast, branching corals like Acropora muricata feature an upright, open structure that promotes high boundary-layer water flow, facilitating particle capture while minimizing the risk of sediment accumulation and algal attachment.15,16 Comparing these two species is ecologically and practically significant: A. hyacinthus is a primary reef-builder with high restoration value but is highly sensitive to sedimentation, whereas A. muricata is a resilient, fast-growing species often used for rapid biomass production. These species serve as ideal models to determine if a universal feeding protocol is feasible or if morphology-specific management is a prerequisite for RAS efficiency.17
This study aimed to evaluate and compare the species-specific responses of Acropora hyacinthus and Acropora muricata micro-fragments to different regimes of organic feeding and trace element supplementation. Understanding these interactions is essential for optimizing land-based coral nurseries, particularly in the South Central Coast of Vietnam, where these species are dominant but face varying environmental pressures. By analyzing survival rate, specific growth rate (SGR), and absolute growth rate (AGR) across different fragment sizes, this study seeks to establish a practical, morphology-driven nutritional framework for the sustainable cultivation of Acropora. The results contribute to optimizing coral production for large-scale restoration while reducing risks associated with nutrient management in recirculating systems.
Materials and Methods
Experimental design and system configuration
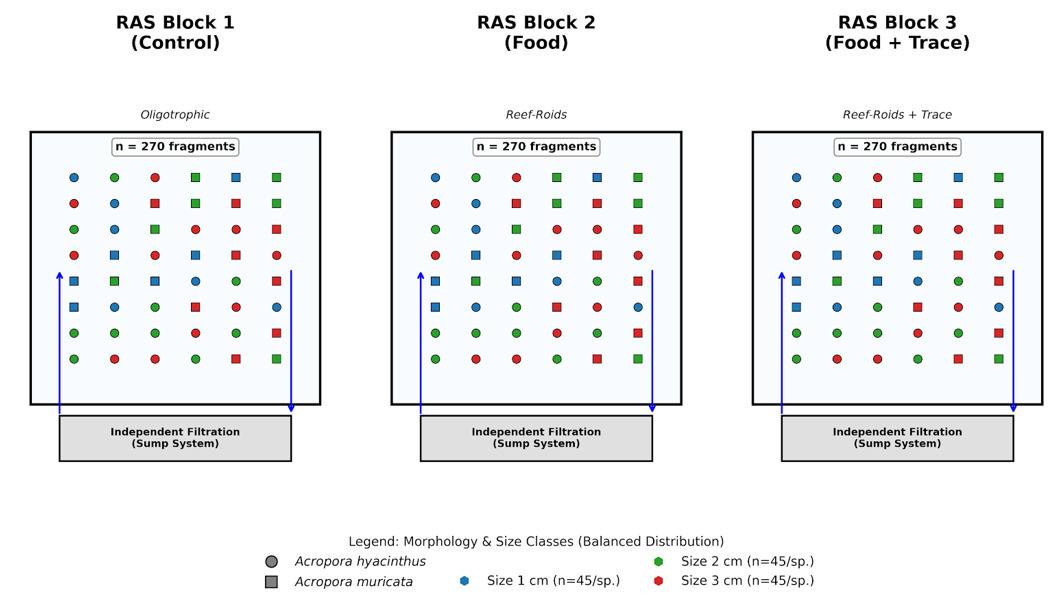
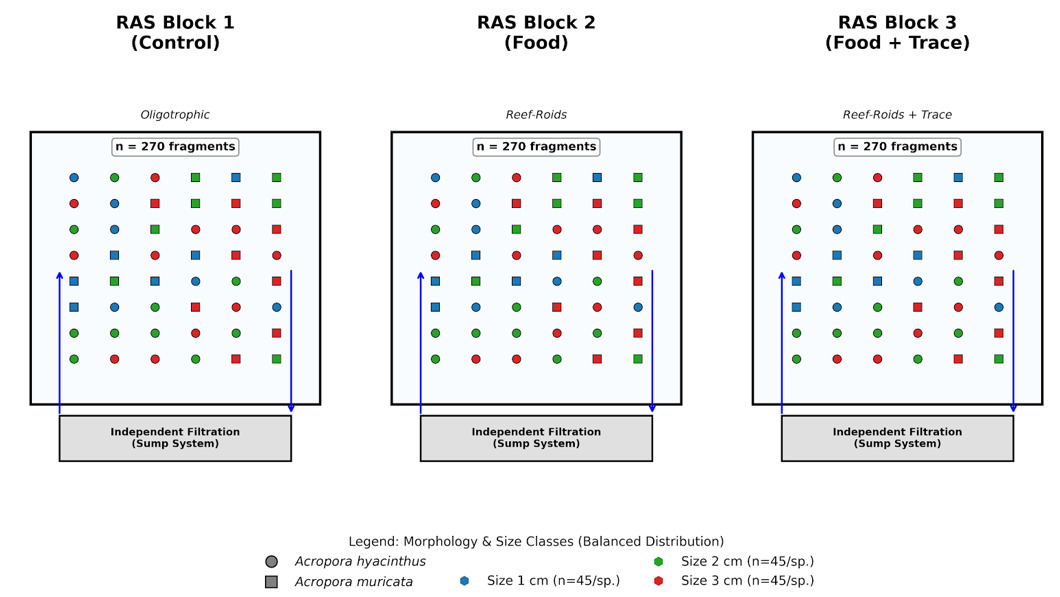
The study was conducted at the coral aquaculture facility of the Coastal Branch, Joint Vietnam-Russia Tropical Science and Technology Research Center. We utilized a controlled experimental design to evaluate the effects of nutritional regimes and initial fragment size on two scleractinian corals: Acropora hyacinthus (tabular) and Acropora muricata (branching). A total of 810 micro-fragments were utilized, with 45 fragments per species per size class allocated across three independent 400-L Recirculating Aquaculture Systems (RAS), each representing a specific nutritional treatment (Table S1, Figure S1). To mitigate potential system-level bias (“tank effect”), environmental parameters were synchronized across all units (Salinity: 34 ± 0.5 ppt; temperature: 26 ± 0.5oC; pH: 8.1 ± 0.1). Lighting was provided by Sea Torch LED (LM 25 Pro) fixtures at a 10-hour photoperiod (150 – 250 µmol photons.m-2.s-1). Within each tank, fragments were randomly assigned to positions on the cultivation racks to eliminate bias from localized flow or light gradients.
Micro-fragmentation and biological management
Mother colonies were fragmented into three size classes (1, 2, and 3 cm) using a high-precision diamond band saw (C-40, Gryphon) and mounted onto ceramic plugs with cyanoacrylate gel. All fragments were acclimated for 14 days prior to the experimental treatments. Fragments were acclimated for 14 days to ensure tissue recovery before the trial. To stabilize inorganic nutrients and suppress filamentous algae, a microbial consortium (MicroBacter7, Brightwell Aquatics; 1 x 106 CFU/mL) was inoculated weekly. A liquid carbon source (Reef BioFuel, Brightwell Aquatics) was dosed daily at 0.5 mL.100 L-1 to maintain the C:N:P balance. Nitrate (NO3-) and phosphate (PO43-) were measured weekly using Marine Master Multiparameter Photometer (Hanna - Hi 97115C, Italia).
Nutritional treatments and adaptive feeding
Fragments were assigned to one of three regimes: (1) Control: oligotrophic conditions without external feeding; (2) Food: particulate organic diet using Reef-Roids (Polyp Lab, USA), administered three times weekly; and (3) Food + Trace: Reef-Roids supplemented with the Balling Light Trace 1-2-3 system (Fauna Marin, Germany). The trace elements (Sr, Ba, I, Fe, and Heavy Metals) were delivered via a Jebao 4.4 automated dosing pump at 0.5 mL.100.L-1 daily. Notably, the feeding dosage was adjusted from 4g (Days 1–45) to 2g (Days 46–88). This adaptive reduction was implemented to optimize the system’s carrying capacity and prevent excessive organic entrapment on the horizontal coenosteum of A. hyacinthus, which was observed to trigger algal encroachment during the initial phase.
Data collection and growth metrics
Survival and growth performance were monitored at regular intervals throughout the 88-day trial. For the tabular Acropora hyacinthus, growth was quantified based on 2D surface area (mm2) derived from top-down digital photography and ImageJ software analysis. For the branching Acropora muricata, growth was measured as linear extension (mm) using high-precision calipers. Two primary growth metrics were calculated to evaluate performance. The Absolute Growth Rate (AGR) was determined to quantify the average daily increment in physical dimensions using the following formula:
AGR=(Mf-Mi)/Δt
where Mf and Mi represent the final and initial measurements (area or length), respectively, and Δt is the duration of the experimental period in days.
To evaluate the relative growth potential independent of initial size, the Specific Growth Rate (SGR) was calculated using the logarithmic formula:
SGR(%⋅day-1)=((lnMf-lnMi)/Δt)×100
Statistical Analysis
Statistical analyses were performed in R (v. 4.5.3). Data were tested for normality using the Shapiro-Wilk test and for homoscedasticity using Levene’s test. A Two-way ANOVA was used to assess the effects of treatment and size class. Post-hoc comparisons were conducted using Tukey’s HSD (p < 0.05). Survival dynamics were analyzed using the Cox Proportional Hazards model, after verifying the proportional hazards assumption via Schoenfeld residuals.
Results
Survival performance and hazard ratio analysis
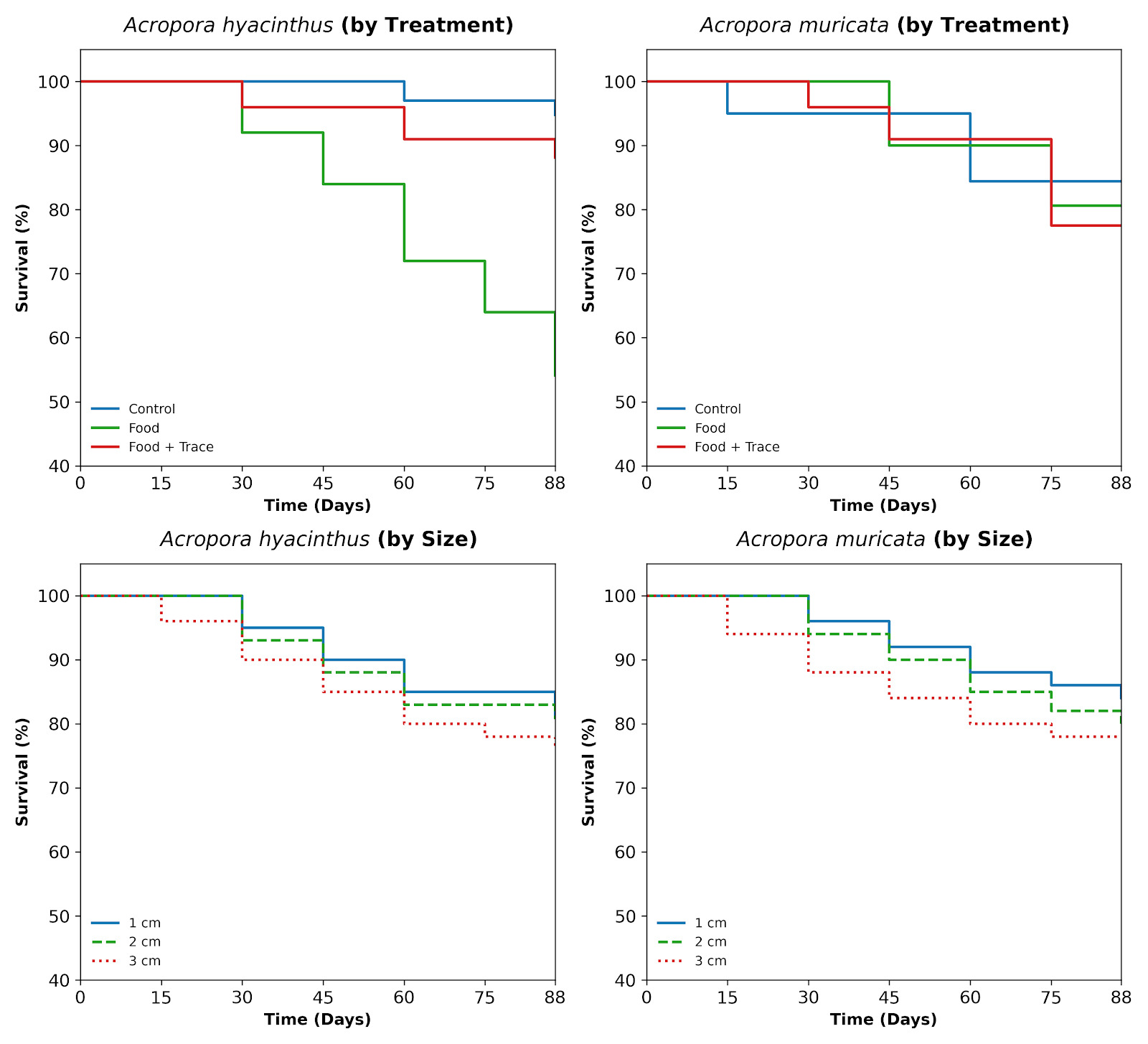
The cumulative survival analysis revealed distinct resilience strategies between the two species under the experimental nutritional regimes (p < 0.001). The tabular species, A. hyacinthus, exhibited extreme sensitivity to organic loading, whereas the branching species, A. muricata, maintained a more stable, albeit declining, survival rate across all treatments. To quantify the relative risk of mortality, a Cox Proportional Hazards model was applied using the “Control” group as the reference (HR = 1.0). For A. hyacinthus, the “Food” treatment resulted in a Hazard Ratio (HR) of 10.22 (p < 0.001), indicating that fragments in this group were ten times more likely to die than those in the control. The “Food + Trace” treatment showed a mitigated risk (HR = 2.35, p = 0.059), but remained significantly higher than the control (Table 1).
The temporal dynamics show a sharp decline in A. hyacinthus survival in the “Food” group after Day 53, whereas A. muricata displayed a more consistent mortality rate throughout the 88-day period (Figure 1). The cumulative survival curves were plotted as a function of the number of days elapsed from the beginning of the experiment (Day 0: 2024-08-05). This visualization clearly demonstrates the temporal vulnerability of the two species across different cultivation phases.
_and_*a._muricata*_(right).png)
For A. hyacinthus, mortality in the Food treatment was relatively moderate until Day 53 (approx. 70% survival), but accelerated significantly during the final phase (Day 53 to Day 88), concluding at 54.1% (Figure 1). This suggests that the physiological stress in tabular corals, likely due to organic matter accumulation and algal competition, reaches a critical “tipping point” after approximately two months of continuous intensive feeding. For A. muricata, the survival trends were more linear. Significant mortality events occurred primarily within the first 2 days and the final 10 days of the 88-day experiment. The branching structure showed a more consistent resilience to nutrient loading compared to the tabular morphology.
Growth performance: Absolute growth rate and specific growth rate
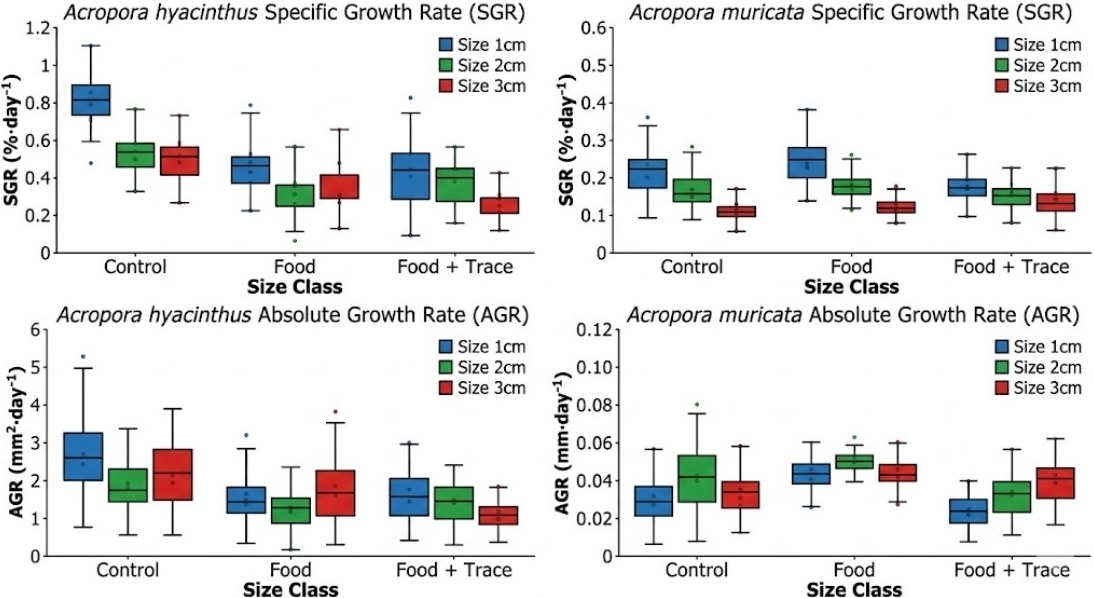
The analysis of growth performance revealed distinct physiological trajectories for A. hyacinthus and A. muricata under the tested nutritional regimes. These differences in growth metrics were closely linked to the interaction between fragment size and each species’ specific colony architecture (Figure 2).
_and_fragment_size_(1__.jpg)
The growth dynamics of the tabular species, A. hyacinthus, revealed a significant sensitivity to organic loading and initial fragment size. The highest specific growth rate (SGR) across all experimental groups was observed in the 1 cm Control group, yielding 0.89 ± 0.37% .day-1 (Table 2, p < 0.05). This group also demonstrated the highest absolute growth rate (AGR) at 3.24 ± 1.25 mm2.day-1, suggesting that under oligotrophic conditions, smaller micro-fragments possess a superior capacity for rapid coenosteum expansion. A distinctive “nutritional paradox” was identified in this species. Contrary to expectations of heterotrophic enhancement, the introduction of the particulate diet (Reef-Roids) resulted in a marked decline in growth performance. For 1 cm fragments, the SGR decreased significantly to 0.51 ± 0.38%. day-1 in the Food treatment. This inhibitory effect was accompanied by a drastic reduction in sample size (n), where the number of surviving fragments in the Food and Food + Trace groups plummeted to as low as n = 20–34, compared to n = 42–44 in the Control. These data indicate that the horizontal architecture of A. hyacinthus may facilitate the detrimental entrapment of organic particles, leading to tissue necrosis and suppressed calcification.
In contrast to the tabular species, the branching A. muricata exhibited a neutral to positive response to heterotrophic supplementation. The maximum SGR was recorded in the 1 cm Reef-Roids treatment (0.27 ± 0.14%.day-1), which slightly exceeded the performance of the corresponding Control group (0.23 ± 0.17%.day-1). Although the overall growth rates of the branching species were lower than those of the tabular species, A. muricata maintained higher physiological stability under high organic loads, with relatively consistent survival counts (n) across all treatments. Linear extension (AGR) for A. muricata reached a steady peak at the 2 cm size class in both Control and Food treatments (0.048 mm.day-1), suggesting that 2 cm may be a more resilient mechanical unit for linear skeletal development in branching forms. However, the SGR remained inversely proportional to fragment size; 3 cm fragments consistently yielded the lowest growth rates (0.11–0.13%.day-1), marked by significant statistical differences (Tukey’s HSD, p < 0.05, superscript ‘b’) relative to the 1 cm cohorts.
The photographic evidence the profound impact of colony morphology on aquaculture success (Figure 3). As visually confirmed, all size classes of A. muricata exhibited robust linear extension and healthy tissue integration, particularly the 1 cm fragments under the Reef-Roids regime (Figure 3B), which achieved the highest species-specific growth. Conversely, the final growth state of A. hyacinthus (Figure 3A) reveals significant tissue recession and algal encroachment in the “Food” treatment groups. This visual divergence provides a biological basis for the “nutritional paradox” identified in the statistical analysis, where the entrapment of organic matter on the horizontal coenosteum of the tabular morphology led to growth inhibition and a tenfold increase in mortality risk reported.
_the_tabular_*acropora_hya.jpeg)
Influence of initial fragment size on re-epithelialization
Across both species and all nutritional regimes, a consistent ontogenetic trend was observed: the 1 cm size class achieved the highest SGR (Figure 2). This phenomenon is likely attributed to the higher surface area-to-volume (S/V) ratio characteristic of smaller fragments, which optimizes metabolic exchange and enhances the regenerative stimulus triggered by the fragmentation process. While the 1 cm fragments exhibited the highest growth potential, their vulnerability in land-based RAS was strictly morphology-dependent. In A. hyacinthus, the enhanced metabolic demand of 1 cm fragments was neutralized by the negative impacts of organic accumulation. Conversely, the open architecture of A. muricata likely promoted superior boundary-layer water flow, preventing particle settlement and allowing the species to harness heterotrophic energy without the associated hazard of organic entrapment. These findings emphasize that for high-performance coral mariculture, size optimization must be integrated with species-specific morphological management.
Statistical interactions of size and nutritional regime
Statistical evaluation using Two-way ANOVA confirmed that initial fragment size was the most significant driver of growth across both species (p < 0.001, Table 3). For A. hyacinthus, the interaction between nutritional treatment and size class was marginally significant (F = 1.98, p = 0.097), suggesting that larger tabular fragments may be more susceptible to the stressors associated with high organic loading. Conversely, A. muricata showed no significant interaction between treatment and size (F = 0.64, p = 0.634), reinforcing the observation that branching structures maintain a consistent growth response across different size classes regardless of Reef-Roids or Balling Light Trace supplementation (Table 3). This lack of interaction emphasizes that while branching corals are structurally robust to particulate feeding, their relative growth potential remains fundamentally constrained by their initial ontogenetic stage.
Discussion
Morphological determinism in particulate heterotrophy
The divergent survival and growth responses observed between A. hyacinthus and A. muricata emphasize that colony architecture is a primary determinant of nutritional success in land-based aquaculture. While heterotrophic feeding is widely recognized as a mechanism to enhance coral calcification and resilience,6 our results demonstrate a “morphological filter” effect. The flat, horizontal coenosteum of the tabular A. hyacinthus acted as a physical trap for the Reef-Roids particulate powder. This accumulation, compounded by a high feeding frequency (3 times weekly), likely created localized hypoxic conditions and promoted the rapid proliferation of filamentous algae.
This result is consistent with previous studies indicating that corals with horizontal growth forms are more vulnerable to sedimentation and particulate stress than branching morphologies.14 The catastrophic mortality risk (HR = 10.22) in the A. hyacinthus Reef-Roids group supports the “microbialization” hypothesis,10 where excessive organic loading on the coral surface shifts the microbial balance toward opportunists, leading to tissue necrosis. Conversely, the vertical orientation and open structure of A. muricata facilitated superior water flow and waste removal, preventing particulate settlement and allowing the coral to maintain stable survival.
Hydrodynamics and system-level interactions in RAS
The species-specific performance observed in this study is inextricably linked to the hydrodynamic conditions within the land-based RAS. In a closed recirculating system, flow dynamics dictate the rate of particle settlement and the thickness of the diffusive boundary layer surrounding the coral tissue. Our results suggest that the horizontal coenosteum of A. hyacinthus created a “stagnant zone,” characterized by reduced water velocity and increased particulate retention. This is consistent with the findings of Anthony and Fabricius,14 who noted that tabular morphologies incur higher metabolic costs in environments with low flow and high organic loads due to the necessity of active sediment rejection via mucus production. In our RAS configuration, although the filtration efficiency was maintained through mechanical and biological sumps, the physical entrapment of Reef-Roids powder on the flat surfaces of A. hyacinthus likely triggered localized microbial shifts. The accumulation of organic matter provides a substrate for opportunistic filamentous algae, which, in the absence of high-velocity “flushing” flows, rapidly smother the coral micro-fragments.11 This explains the significant tissue recession and subsequent high mortality observed in the fed tabular groups. Conversely, the upright architecture of A. muricata promotes turbulent mixing, minimizing the boundary layer thickness and facilitating the removal of metabolic waste. Furthermore, the environmental homogeneity across the independent RAS units ensures that the observed differences are treatment-driven rather than due to tank-specific confounding factors.
The nutritional paradox: Growth inhibition vs. caloric gain
The growth performance metrics revealed a significant nutritional paradox for the tabular morphology. Although heterotrophy can provide up to 100% of the daily metabolic requirements for some corals.7 A. hyacinthus achieved its maximum SGR (0.89%.day-1) and AGR (2.59 mm2.day-1) under oligotrophic Control conditions. The reduction in growth when fed Reef-Roids suggests that for tabular species, the metabolic energy diverted toward “cleaning” mechanisms—such as mucus production and ciliary action to remove particulate matter—exceeded the nutritional energy gained from the food.
In contrast, the 1 cm micro-fragments of A. muricata exhibited a positive response to heterotrophy (SGR 0.27.day-1), confirming that branching structures are better adapted to leverage particulate organic matter for linear extension. This pattern agrees with previous work indicating that horizontal growth forms incur higher metabolic costs in nutrient-rich environments, largely due to greater exposure to algal competition and sediment accumulation.17
Synergistic effects of trace elements and micro-fragmentation
A novel aspect of this study was the mitigating effect of Fauna Marin Balling Light Trace 1-2-3 on mortality. The systematic delivery of essential ions (Strontium, Barium, Iodine, and Heavy Metals) via automated dosing significantly improved the survival of A. hyacinthus from 54.1% (Food only) to 88.1% (Food + Trace). This suggests that trace elements act as vital physiological buffers, supporting enzymatic functions and skeletal mineralization under high-stress organic loading.3 However, the fact that growth still lagged behind the Control group indicates that chemical supplementation can support maintenance and survival but cannot fully compensate for the physical stressors of bio-fouling on horizontal surfaces.
Furthermore, the superior relative growth (SGR) of 1 cm micro-fragments across both species reinforces the “edge effect”.6 Smaller fragments have a higher peripheral-to-surface area ratio, which promotes rapid calcification during the healing process. Our results confirm that micro-fragmentation is a highly efficient strategy for rapid biomass production, provided that the environmental parameters and nutritional regimes are tailored to the specific morphology of the target species.
Practical implications and conclusions
The Hazard Ratio and growth analyses provide actionable guidelines for coral nursery management. For tabular species like A. hyacinthus, practitioners must prioritize oligotrophic conditions and high water turnover to prevent catastrophic mortality. Conversely, branching A. muricata can leverage intensive heterotrophic feeding to maximize linear extension. Managing nutrients through microbial consortia and carbon dosing remains essential for maintaining water quality in fed systems. We propose a stratified management protocol: a “low-nutrient” track for horizontal morphologies and a “high-nutrient” track for branching forms to optimize RAS production efficiency. This 88-day study confirms that colony architecture dictates the success of nutritional supplementation. While heterotrophy is a viable energy-enhancement strategy, its application is strictly species-specific. A. hyacinthus is highly vulnerable to organic loading; its horizontal architecture traps debris and promotes algal proliferation, increasing mortality risk tenfold (HR = 10.22). In contrast, A. muricata exhibits superior resilience, as its upright structure minimizes bio-fouling and facilitates efficient nutrient uptake. Across both species, the 1 cm size class consistently showed the highest relative growth potential, making it the optimal biological unit for rapid biomass production. While trace element supplementation serves as a vital physiological buffer for tabular species under organic stress, it cannot fully compensate for the growth-inhibiting effects of algal competition. Ultimately, land-based restoration programs must adopt morphology-specific management to maximize overall nursery efficiency.
Acknowledgments
We thank the Coastal Branch of the Joint Vietnam-Russia Tropical Science and Technology Research Center (VB.Đ1.08/23) for providing facilities and technical support. Gratitude is extended to the staff for their assistance in RAS maintenance and data acquisition.
Authors’ Contribution - CRediT
Conceptualization: Viet D.H. Dang (Lead). Methodology: Viet D.H. Dang (Equal), Hai-Thanh T. Nguyen (Equal). Formal Analysis: Viet D.H. Dang (Equal), Thuy T.P. Dinh (Equal), Thanh V. Dang (Equal), Nghi Q. Pham (Equal), Hanh D. Vu (Equal). Investigation: Viet D.H. Dang (Equal), Thuy T.P. Dinh (Equal), Thanh V. Dang (Equal), Nghi Q. Pham (Equal), Hanh D. Vu (Equal). Writing – review & editing: Viet D.H. Dang (Equal), Hai-Thanh T. Nguyen (Equal), Dien D. Tran (Equal), Ha T. Vo (Equal), Lam N. Hoang (Equal), Lien P. Nguyen (Equal).
Ethical conduct approval – IACUC
Not required.
Competing of Interest – COPE
No competing interests were disclosed.
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.