Introduction
Aquaculture is one of the fastest-growing sectors of global food production and plays a crucial role in food security and socioeconomic development, particularly in developing regions.1–3 Among cultured species, the bighead catfish (C. macrocephalus) is widely farmed in freshwater areas in Southeast Asia due to its high market value and strong environmental tolerance. This species possesses accessory respiratory organs that facilitate aerial respiration and survival in hypoxic environments.4,5 Recent findings have also indicated that bighead catfish can perform well under low-salinity conditions (2‰), highlighting its potential for aquaculture in regions increasingly affected by salinity intrusion.6
Salinity intrusion driven by climate change has become an increasing challenge in aquaculture, particularly in low-lying coastal and delta regions such as the Mekong Delta of Vietnam and other parts of Southeast Asia, adversely affecting agriculture, fisheries, and biodiversity. Even slight increases in salinity can disrupt osmotic and ionic balance and elevate osmoregulatory energy demands. These changes may induce oxidative stress, impair immune function, increase disease susceptibility, and reduce growth performance and survival in freshwater fish species. These effects ultimately lead to reduced productivity and higher production costs for farmers. These impacts are further exacerbated in intensive aquaculture systems by additional stressors, including high stocking densities and fluctuating water quality.7,8 Although antibiotics are commonly used to mitigate such challenges in farmed fish, their overuse has raised serious concerns regarding antimicrobial resistance, environmental contamination, and food safety.9,10 Consequently, there is growing interest in sustainable alternatives such as phytogenic feed additives.11,12
Garlic (Allium sativum) is an inexpensive and readily available phytogenic feed additive. In particular, it is rich in bioactive organosulfur compounds, particularly allicin, which exhibit antimicrobial, antioxidant, and immunomodulatory properties.13,14 Garlic has been reported to support the health and welfare of farmed species through multiple biological pathways, including modulation of gut microbiota, stimulation of digestive enzyme activity, and improved nutrient utilization.15,16 It also strengthens antioxidant defenses by scavenging reactive oxygen species and stimulating endogenous antioxidant systems, thereby helping to maintain redox balance under stress conditions.17,18 Garlic application via feed or immersion can enhance the performance of cultured aquatic species. Its effects depend on garlic form, dosage, species, life stage, and duration of application.10 Dietary garlic powder has been tested across a wide range (0.05–40 g kg⁻¹),19 with optimal responses commonly reported within the range of 10–20 g kg⁻¹.19–22 However, higher levels show inconsistent effects across species. For example, 40 g kg⁻¹ improved growth performance and feed utilization in Lates calcarifer,23 whereas 25 g kg⁻¹ reduced performance in Lateolabrax japonicus,20 and levels above 20 g kg⁻¹ adversely affected red blood cell indices and hematocrit in Dicentrarchus labrax.24
Under salinity stress, fish often exhibit altered metabolic and physiological states. These changes are frequently accompanied by variations in hematological parameters, which serve as indicators of health status and stress response.25–28 In scaleless catfish such as the bighead catfish, the absence of protective scales may increase sensitivity to handling and environmental stressors.29–31 This can lead to more pronounced alterations in their metabolic and physiological states under salinity stress. Although there is considerable evidence regarding the effects of dietary garlic on the performance of farmed fish, literature associated with its influence on C. macrocephalus under low-salinity conditions remains limited. This knowledge gap is particularly relevant given the increasing prevalence of salinity intrusion into freshwater aquaculture systems and the species’ potential for culture in such environments. Therefore, this study evaluated the effects of dietary garlic supplementation (0–30 g kg⁻¹) on growth performance, survival, and hematological responses of juvenile bighead catfish under low-salinity (2‰) conditions. The study aims to provide practical insights to support the sustainable culture of this species in salinity intrusion–affected regions.
Materials and Methods
Experimental fish and acclimation
Juvenile bighead catfish with an average body weight of 9.61 ± 1.62 g and total length of 10.42 ± 1.16 cm were obtained from a local hatchery and transported to the freshwater aquaculture experimental facility at Tra Vinh University. Transportation followed the recommended live fish-handling procedures.32 After arrival, fish were acclimated for one week in 1 m³ aerated composite tanks. Initially, the fish were maintained in freshwater (0‰ salinity) and gradually acclimated to 2‰ salinity by increasing salinity by 1‰ every 12 h using 30‰ bittern water collected from nearby salt fields. After reaching the target salinity, fish were maintained under the same conditions for an additional 3 days prior to the start of the feeding trial. During acclimation, fish were fed a basic commercial diet once daily. Uneaten feed and accumulated waste were removed daily to maintain suitable water quality. Throughout the acclimation period, fish exhibited normal swimming and feeding behavior.
Freshwater used in the experiment was collected from a nearby river, allowed to settle to remove suspended particles, disinfected with potassium permanganate (KMnO₄) at 5 mg L⁻¹, and aerated for three days before use. The bittern water (30‰) was treated with chlorine at 40 mg L⁻¹ and aerated for three days. Freshwater and saltwater were mixed to obtain the desired salinity of 2‰ using the dilution equation C₁V₁ = C₂V₂, where C₁ represents the salinity of the saltwater, V₁ is the volume of saltwater used, C₂ is the target salinity, and V₂ is the total volume of the mixture.
Diet preparation
Commercial pellets containing 30% crude protein (DACHAN Joint Stock Company, Vietnam) served as the basal diet. The proximate nutritional components of the basal feed are detailed in Table 1. Garlic cloves were sourced from a local market, peeled, and then oven-dried at 70°C for 13 hours. Both the dried garlic and the commercial pellets were ground separately into fine powders, sealed in polyethylene bags, and stored at 4°C until needed. Ten minutes before feeding, the required amounts of garlic powder were thoroughly suspended in 1 mL of water. The solutions were then thoroughly mixed with the basal feed powder to obtain levels (0, 10, 20, and 30 g kg⁻¹).
Experimental design and feeding management
This study was conducted at the Freshwater Aquaculture Experimental Hatchery of Tra Vinh University (Vietnam) from 30 July to 30 October 2025. The experimental design, including the sample size and replication level, was consistent with established practices in aquaculture nutrition studies evaluating phytogenic feed additives. A completely randomized design was employed with four dietary treatments (0, 10, 20, and 30 g kg⁻¹ garlic powder), each in triplicate. A total of 240 fish were randomly assigned to 12 aerated composite tanks (250 L capacity; 100 L working volume) at a stocking density of 20 fish per tank. The culture system was operated as a semi-static system with continuous aeration and periodic water exchange, in which approximately 30% of the water volume in each tank was replaced weekly. The feeding trial lasted 60 days under conditions of 2‰ salinity. Fish were fed twice daily (07:30 and 16:00) at 2–3% of their body weight. Feed intake was monitored in each tank at every feeding, and amounts were adjusted based on previous feeding activity. No abnormal feeding behavior was observed among treatments throughout the trial. Uneaten feed and feces were removed daily, and approximately 30% of the water was replaced weekly.
Growth performance measurements
Fish were measured at 15-day intervals throughout the experiment. At each sampling point, five fish per tank were randomly selected for measurement. Fish were gently blotted dry and weighed using an electronic balance (±0.01 g), and total length was measured to the nearest millimeter. Survival, productivity, and feed conversion ratio were recorded at the end of the experiment. Monitored performance parameters were calculated using standard formulas commonly applied in aquaculture studies.
Mean weight (MW, g) = total weight of sampled fish/number of fish
Mean length (ML, cm) = total length of sampled fish/number of fish
Weight gain (WG, g) = final weight − initial weight
Length gain (LG, cm) = final length − initial length
Daily weight gain (DWG, g day⁻¹) = (final weight − initial weight) / rearing days
Daily length gain (DLG, cm day⁻¹) = (final length − initial length) / rearing days
Specific growth rate in weight (SGRW, % day⁻¹) = [(ln final weight − ln initial weight) / rearing days] × 100
Specific growth rate in length (SGRL, % day⁻¹) = [(ln final length − ln initial length) / rearing days] × 100
Survival rate (%) = (final fish number / initial fish number) × 100
Productivity (g m⁻3) = total biomass / rearing volume
Feed conversion ratio (FCR) = total dry feed fed (g) / total wet weight gain (g)
Hematological analysis
At the end of the experiment, three fish per tank (nine fish per treatment) were randomly selected for hematological analysis. Fish were anesthetized using cold shock (ice immersion) prior to blood collection. Blood samples (~0.4 mL) were collected from the caudal vein using sterile syringes and transferred into EDTA-coated tubes. Hematological parameters, including leukocyte (WBC), lymphocyte (LYM), mid-sized cell (MID), and erythrocyte (RBC) counts, were analyzed using standard methods.33,34
Water quality monitoring
Water quality parameters were monitored throughout the experimental period. Temperature and pH were measured twice daily using a portable meter (HANNA HI98103, Romania), while salinity was measured using a salinometer (YSI, Japan). Total ammonia nitrogen (TAN) and nitrite (NO₂⁻) were measured every three days using commercial test kits (Sera, Germany).
Data analysis
Statistical analyses were conducted using one-way analysis of variance (ANOVA), followed by Duncan’s multiple range test to compare treatment means at a significance level of P ≤ 0.05. Before analysis, data were assessed for normality and homogeneity of variances. Percentage data were arcsine-transformed when necessary to satisfy ANOVA assumptions. All analyses were performed using IBM SPSS Statistics for Windows (version 20.0).
Results
Water quality parameters
Water quality remained stable throughout the 60-day experimental period, with only minor variation among treatments (Table 2). Water temperature ranged from 27.74 to 28.88°C, and pH values were maintained between 6.92 and 7.26. TAN and NO₂⁻ concentrations remained low across all treatments, ranging from 0.01 to 0.02 mg L⁻¹ and 0.01 to 0.03 mg L⁻¹, respectively.
Growth performance and hematological responses
Growth performance in weight showed a dose-dependent pattern across dietary treatments. At the end of the experiment, fish fed the control (0 g kg⁻¹) and 20 g kg⁻¹ garlic diets exhibited similar values of final mean weight (FMW), WG, DWG, and SGRW, which were significantly higher than those of fish fed 10 g kg⁻¹ (P ≤ 0.05, Table 3). Fish fed 30 g kg⁻¹ showed intermediate values and did not differ significantly from the other treatments (P ≥ 0.05, Table 3).
Length-related parameters were generally less responsive to dietary treatments. At day 45, ML₄₅ was significantly lower in fish fed 10 g kg⁻¹ compared with the other treatments (P ≤ 0.05, Table 4). However, no significant differences were observed among treatments for FML, LG, DLG, or SGRL (P ≥ 0.05, Table 4).
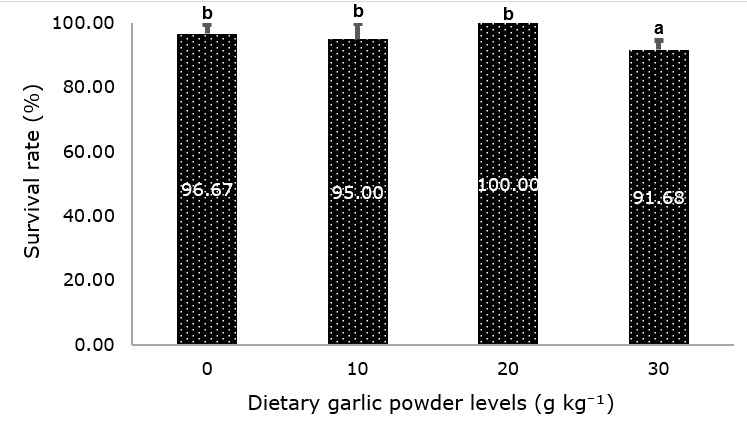
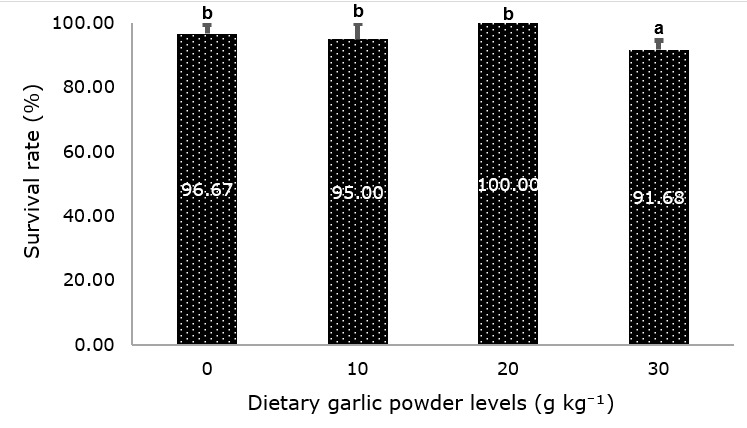
The highest survival (100%) was observed in fish fed 20 g kg⁻¹ garlic, while survival remained high (95–97%) in the control and 10 g kg⁻¹ treatments, with no significant differences (P ≥ 0.05, Figure 1). In contrast, fish fed 30 g kg⁻¹ exhibited a significantly lower survival rate than the other treatments (P ≤ 0.05, Figure 1). Productivity and FCR were not significantly affected by dietary treatments (Table 5, P ≥ 0.05).
WBC counts were highest in fish fed 20 g kg⁻¹ garlic and were significantly higher than those in fish fed 30 g kg⁻¹ (P ≤ 0.05; Table 6) but did not differ significantly from the control or 10 g kg⁻¹ treatments (P ≥ 0.05; Table 6). No significant differences were observed among treatments for LYM, MID, or RBC counts (P ≥ 0.05; Table 6).
Overall, the results consistently demonstrated a non-linear dose–response pattern across growth in weight, survival, and hematological parameters.
Discussion
Growth performance
This study evaluated the effects of dietary garlic powder supplementation on the growth performance of juvenile C. macrocephalus under low-salinity (2‰) conditions. Fish fed 20 g kg⁻¹ garlic exhibited growth performance in weight (via FMW, WG, DWG, and SGRW) comparable to the control and significantly higher than that of fish fed 10 g kg⁻¹, whereas the 30 g kg⁻¹ treatment produced intermediate results without significant differences among other treatments. These findings indicate that 20 g kg⁻¹ represents an effective supplementation level for maintaining stable growth. In contrast, both lower and higher supplementation levels showed no additional benefit and a tendency toward reduced performance under the present conditions.
The significantly reduced growth performance observed at 10 g kg⁻¹ may be attributed to an insufficient level of bioactive compounds in dietary garlic to elicit beneficial physiological effects. Although C. macrocephalus can tolerate low salinity (2‰),6 maintaining osmotic and ionic balance in such environments requires additional metabolic energy.35,36 This increased demand may elevate metabolic load and disrupt redox balance,8 thereby reducing energy available for growth and immune function. Garlic-derived bioactive compounds, particularly organosulfur compounds such as allicin, may help mitigate these effects by enhancing digestive efficiency, improving gut microbial balance, and strengthening antioxidant defenses.9,16,17 However, these benefits are dose-dependent, and insufficient supplementation may fail to activate these mechanisms effectively.37,38 These can shift energy allocation away from somatic growth toward maintenance functions.8,39 Moreover, low doses may impose additional metabolic costs associated with physiological adaptation to phytogenic compounds without providing compensatory benefits. As a result, these negative effects may outweigh potential advantages, leading to unchanged or slightly reduced growth performance.15,40
On the other hand, the reduced growth performance trend and significantly lower survival observed at 30 g kg⁻¹ may be attributed to the adverse effects of excessive garlic supplementation. While bioactive compounds in garlic have been reported to support gut microbiota balance, digestive efficiency, and nutrient absorption9,21,41 and have immunomodulatory, antioxidant, anti-inflammatory, and antibacterial features,42,43 excessive levels may disrupt physiological homeostasis. High levels of garlic-derived organosulfur compounds, particularly allicin, may exert cytotoxic and pro-oxidant effects, leading to oxidative stress, lipid peroxidation, and tissue damage. In addition, excessive garlic intake may cause intestinal mucosal damage, alter gut microbial balance, and induce gastrointestinal irritation and metabolic stress, as reported in Oreochromis niloticus, Cyprinus carpio, and Perca fluviatilis.21,44,45 Furthermore, high inclusion levels have been associated with hematological disturbances, including erythrocyte damage and reduced oxygen-carrying capacity, as well as hepatic dysfunction, indicating increased metabolic and detoxification burdens.38,46 The strong odor and pungency of garlic at excessive levels may also reduce feed palatability and intake, thereby limiting nutrient assimilation and growth.38,44,47 Collectively, these disturbances can redirect energy allocation toward stress responses, detoxification, and cellular repair rather than somatic growth, ultimately resulting in decreased growth performance and survival.
Optimal dietary garlic supplementation levels may enhance physiological stability and energy allocation toward growth and survival.17,18 In the present study, growth performance did not differ significantly from the control, while the highest survival rate was observed at the 20 g kg⁻¹ supplementation level, indicating that the fish were already approaching their growth potential under the experimental conditions. These findings indicate that dietary garlic may contribute more to homeostasis, stress tolerance, and immune function than to growth promotion under low-salinity conditions. These explanations are consistent with the lack of significant differences in FCR and productivity among treatments. Similar responses have also been reported in several species.47,48
Another result showed that dietary garlic treatments did not significantly affect length growth parameters, despite variations in weight-based parameters. This divergence likely reflects differences in the regulation of somatic and skeletal growth in fish. Body weight responds rapidly to changes in nutrient intake and energy allocation, whereas length growth is more tightly controlled by endocrine mechanisms and structural processes such as bone formation.39,49–51 Consequently, dietary interventions may influence tissue deposition without producing proportional changes in length over relatively short experimental periods. Similar patterns have been reported in Nile tilapia and African catfish, where weight gain responded more readily than length growth to dietary garlic supplementation.52,53
Hematological responses
Monitored hematological parameters remained within the normal ranges reported for Clarias species,26,27,54–56 suggesting that garlic supplementation regulated, rather than disrupted, the physiological status of the experimental juveniles.
A significant effect was observed in WBC counts, which were highest at 20 g kg⁻¹. This increase suggests a stimulation of immune-related processes. WBCs play a central role in innate immunity, including pathogen recognition and inflammatory responses.57–59 Garlic-derived organosulfur compounds, particularly allicin, have been reported to enhance WBC activity, modulate immune signaling pathways, and improve antioxidant defenses.17,38,60,61 Therefore, elevated WBC levels at the 20 g kg⁻¹ supplementation level may reflect enhanced immune readiness, which is consistent with the highest survival observed in this group. In contrast, reduced WBC counts at 30 g kg⁻¹, together with a significant decline in survival rate, may indicate that excessive garlic inclusion adversely affects physiological balance or immune efficiency. This supports a dose-dependent (hormetic) response.62,63 Comparable dose-dependent responses to garlic supplementation have been reported in several fish species.44,47,64 The absence of significant changes in LYM, MID, and RBC suggests that the hematological response was selective rather than indicative of broad alterations in blood composition.
During the experiment, key water quality parameters remained stable and within optimal ranges for bighead catfish,65,66 indicating that the observed effects were primarily attributable to dietary treatments. Although garlic supplementation did not significantly enhance growth performance or feed efficiency, the 20 g kg⁻¹ level improved survival and supported physiological stability under low-salinity conditions. These findings suggest that garlic functions primarily as a health-promoting additive, enhancing stress tolerance rather than directly stimulating growth. From a practical aspect, this increase in survival may potentially improve production efficiency and lower the cost per unit of biomass. Given its low prices and availability, garlic use as a feed additive may represent a cost-effective strategy under the study conditions, despite further cost-benefit and field-scale evaluations being required.
In conclusion, this study indicates that a dietary garlic supplementation level of 20 g kg⁻¹ appears to be optimal for juvenile C. macrocephalus reared in low-salinity (2‰) conditions. This supplementation level was effective in maintaining growth performance, supporting survival, and selectively modulating leukocyte (WBC) response. However, this study has several limitations. The lack of tests on digestive enzyme activity, gut microbiota, and functional immune responses limits mechanistic interpretation. Future studies should incorporate these aspects, along with long-term and field-based evaluations, including economic efficiency, to better elucidate the physiological effects and confirm the applicability of garlic supplementation under commercial conditions.
Acknowledgements
We acknowledge the support of time and facilities from Tra Vinh University (TVU) and Ton Duc Thang University for this study.
Author Contributions
Conceptualization: Huynh Kim Huong (Equal), Cheng-Ting Huang (Equal). Methodology: Huynh Kim Huong (Equal), Cheng-Ting Huang (Equal), Doan Xuan Diep (Equal). Formal analysis: Huynh Kim Huong (Equal), Cheng-Ting Huang (Equal). Investigation: Huynh Kim Huong (Equal), Doan Xuan Diep (Equal). Writing—original draft: Cheng-Ting Huang (Equal), Huynh Kim Huong (Equal), Doan Xuan Diep (Equal). Writing—review & editing: Doan Xuan Diep (Lead). Supervision: Huynh Kim Huong (Equal), Cheng-Ting Huang (Equal).
Conflicts of Interest
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
All experimental procedures complied with institutional and national guidelines for the care and use of aquatic animals. Juvenile bighead catfish were anesthetized with ice during weighing, measuring, and blood sampling to minimize harm. After the experiment, fish were returned to holding tanks for further research.
Data Availability
All data are available upon reasonable request.