1. Introduction
Whitmania pigra is listed in the Chinese Pharmacopoeia (2025 Edition) as one of the original animal sources of the medicinal leech. It plays an irreplaceable role in the clinical treatment of cardiovascular diseases such as myocardial infarction and cerebral thrombosis. As the primary species of artificially farmed leech in China.1,2 With the continuous surge in market demand for medicinal leech materials, the wild resources of W. pigra have been declining annually due to overfishing and habitat destruction, and large-scale artificial aquaculture has become the core approach to fill the resource gap. However, with higher cultivation density and greater complexity of the aquaculture environment, disease issues in W. pigra have become increasingly prominent. Among these, erysipelas caused by Aeromonas allosaccharophila is characterized by an acute onset,3 rapid transmission and high mortality. This has become a critical bottleneck restricting the healthy development of the artificial W. pigra aquaculture industry, inflicting severe economic losses on farmers and indirectly compromising the stable supply of medicinal leech resources.
Intestinal probiotics can not only promote the digestion and absorption of nutrients and energy substances in aquatic animals but also enhance their immune functions.4,5 Together, the intestines of aquatic animals and their symbiotic flora form a biochemical barrier against pathogenic attacks and environmental stresses, playing a crucial role in the host’s immune function. An imbalance in the intestinal microbiota can trigger various diseases.6 In Tachysurus fulvidraco suffering from intestinal septicemia, the intestinal microbiota diversity decreased significantly, with a marked enrichment of the pathogenic bacterium Edwardsiella ictaluri and a reduction in the abundance of probiotics such as Cetobacterium.7 Similar trends of intestinal microbial changes have also been observed in diseased aquatic animals, including Penaeus monodon,8 Paralichthys olivaceus,9 and Cyprinus carpio.10 Unlike vertebrate fish that possess both innate and adaptive immunity, the invertebrate annelid W. pigra relies entirely on the innate immune system.11,12 Given the absence of adaptive immunity, all immunomodulators applied to W. pigra exert their effects by boosting components of innate immunity, rather than via lymphocyte-dependent memory responses.
Astragalus polysaccharide, one of the main extracts of Astragalus membranaceus, has significant biological activities, including immunomodulation, anti-inflammatory, and antioxidative effects.13 As a feed additive, APS can promote intestinal development, regulate the balance of the intestinal microbiota, improve the immune performance of the organism, and further inhibit the growth of pathogenic microorganisms.14 APS has been shown to improve the growth performance of Macrobrachium rosenbergii and enhance its innate immune responses as well as resistance against Vibrio anguillarum.15 APS significantly reduces the incidence of Vibrio alginolyticus infection in Larimichthys crocea cultures.16 APS improves growth performance by enhancing antioxidant function, increasing nonspecific immunity, regulating the Toll/NF-κB pathway and the proPO system, and optimizing the intestinal microbiota of Eriocheir sinensis.17 APS has also been applied to regulate the intestinal microbiota and prevent pathogenic infections in the artificial cultivation of various aquatic animals, including Epinephelus fuscoguttatus,18 Oreochromis niloticus,19 and Micropterus salmoides.20 Intramuscular injection was adopted in this study to ensure accurate dose administration and synchronized immune stimulation, thereby eliminating confounding interference from variations in food intake during W. pigra feeding behavior. This approach conforms to established protocols for parenteral administration in aquatic animals, and the injection route is widely utilized in applied fundamental research.21,22
In the artificial cultivation of W. pigra, frequent disease outbreaks often result in large-scale mortality, severely hindering the sustainable development of this industry. Among these diseases, “erysipelas”, which predominantly occurs during the high-temperature period from mid-to-late August every year, is particularly prominent. Infected individuals exhibit symptoms, including muscle rigidity in the clitellar region, body-surface erythema and swelling, and coelomic effusion, and are associated with high mortality rates, thus becoming a key limiting factor in aquaculture production. Previous studies have demonstrated that APS can promote the growth of W. pigra and enhance its immunity and tolerance to heat stress23; however, systematic research on the specific preventive and therapeutic effects of this polysaccharide against erysipelas, as well as on whether it affects host immune mechanisms by regulating the intestinal microbiota, is still lacking. In this study, 16S rDNA amplicon sequencing was used to systematically analyze the effects of Astragalus polysaccharide on the prevention and control of “erysipelas” and the intestinal microbial community of W. pigra, aiming to reveal its potential mechanism of action and provide new theoretical basis and practical guidance for the prevention and treatment of erysipelas.
2. Materials and methods
2.1. Reagents and preparation
Astragalus polysaccharide with a purity of ≥ 90.0% and a molecular weight of 254.69 Da was purchased from Beijing Solarbio Science & Technology Co., Ltd. It is composed of hexuronic acid, glucose, fructose, rhamnose, arabinose, galacturonic acid, and glucuronic acid. The contents of water, ash, and lead were all less than 10%. Sterile distilled water was used as the solvent for APS injection at a concentration of 3.0 mg·mL⁻¹.
2.2. Experimental animals and experimental design
The W. pigra used in this study were obtained from an ecological leech farm in Xinghua County, Jiangsu Province, China. A total of 120 individuals with a body weight of 2.5 ± 0.5 g were used. Species identification was performed according to the morphological description in Fauna Sinica: Hirudinea, and was further confirmed by COI gene sequencing.24 All individuals were verified as W. pigra. Healthy leeches were randomly assigned to 6 aquariums with dimensions of 30 cm (L)×30 cm (W)×30 cm (H), with 20 leeches per aquarium. The leeches were divided into a treatment group and a control group, with 3 biological replicates per group.
In the 6 aquariums designated for sequencing analysis, each leech in the treatment group was intramuscularly injected with 0.4 mL of APS solution at a dose of 0.02 mg/g body weight,21 whereas each leech in the control group was intramuscularly injected with 0.4 mL of sterile distilled water as a placebo. Aeromonas allosaccharophila, a pathogenic bacterium previously isolated and identified in our laboratory, was activated and cultured. The bacterial suspension was serially diluted with sterile water to a final concentration of 1.0×10⁸ CFU/mL. W. pigra in both control and treatment groups were subjected to an immersion challenge using a bacterial suspension of 1.0×10⁸ CFU/mL. During the experiment, the health status of the leeches was observed and recorded daily. Leeches suffering from erysipelas presented with abdominal congestion and hemorrhage, decreased body elasticity, sucker swelling, and reduced adhesion ability. As the disease progresses, severe rotting of the head and tail, coelomic effusion, and clitellum redness and swelling occur.3 In the treatment group, diseased and dead individuals were labeled AinfI, and surviving individuals were labeled AnorI; in the control group, diseased and dead individuals were labeled infI, and surviving individuals were labeled norI. Moreover, aquaculture water samples were collected and labeled InfW. Healthy leeches without APS injection and bacterial challenge were used as blank controls and labeled CK. The number of dead leeches in each of the 6 groups was recorded, and the survival rate (SR) was calculated via the following formula: SR (%) = 100× (Total number - Number of deaths)/Total number.
During the culture period, the leeches were fed river snails twice a day. Continuous aeration was maintained, and the water was changed daily. The water temperature was maintained at 26.86 ± 1.65 °C, measured at 8:00 and 17:00 each day. Water quality was monitored every 3 days to maintain the following conditions: pH 6.50–7.50, hydrogen sulfide (H₂S) concentration < 0.01 mg/L, ammonia (NH₃) concentration < 0.04 mg/L, and dissolved oxygen (DO) concentration > 2.0 mg/L.
2.3. Sample collection
Intestinal samples were collected at the end of the experiment (Day 6). New gloves were worn for each sample collection to avoid cross-contamination. Leeches were rinsed with sterile autoclaved distilled water, anesthetized on ice, and dissected using sterile instruments. Intestinal tissues and their contents were rapidly collected, placed separately into individual 5 mL sterile cryogenic vials, and immediately frozen in liquid nitrogen. All samples were stored at -80 ℃ in an ultra-low temperature freezer until subsequent analysis.
2.4. Intestinal microbiota analysis
2.4.1. Genomic DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from intestinal samples via the SDS method. The quality and concentration of the extracted DNA were detected via 1% agarose gel electrophoresis and a nucleic acid and protein analyzer (Implen GmbH, Munich, Germany), and the DNA was diluted to 1 ng·μL⁻¹ with sterile water. A total of 30 qualified genomic DNA samples, distributed across 5 groups, were sent to Beijing Novogene Co., Ltd. for PCR amplification. The V3–V4 hypervariable regions of the 16S rDNA were amplified via bacterial universal primer pairs with barcodes (341F: CCTAYGGGRBGCASCAG; 806R: GGACTACNNGGGTATCTAAT). After the PCR products were detected by 2% agarose gel electrophoresis, equal volumes were pooled and purified using the Qiagen Gel Extraction Kit (Qiagen GmbH, Hilden, Germany). Library construction was performed with the NEBNext® Ultra™ II DNA Library Prep Kit (New England Biolabs Inc., Ipswich, MA, USA), and the library quality was evaluated via a Qubit® 2.0 fluorometer (Thermo Fisher Scientific, Waltham, MA, USA) and an Agilent Bioanalyzer 2100 System (Agilent Technologies, Inc., Santa Clara, CA, USA). Finally, paired-end sequencing was conducted on the Illumina NovaSeq 6000 platform (Illumina, Inc., San Diego, CA, USA).
2.4.2. Bioinformatics analysis
Paired-end sequencing reads were assigned to their corresponding samples using barcodes, and the barcodes and primer sequences were trimmed. The reads for each sample were assembled using FLASH (V1.2.11) to generate raw tags.25 Quality control was performed with fastp (V0.20.0) software to obtain clean tags. Clean tags were aligned against the Silva 138.1 database via Vsearch (V2.15.0), and chimeras were removed to yield effective tags.26 Denoising and filtering were performed using the DADA2 module in QIIME2 (Version QIIME2-202006) to obtain the final amplicon sequence variants (ASVs) and feature tables.27,28 Finally, the ASVs were aligned against the reference database via QIIME2 software to obtain taxonomic information for each ASV, followed by multiple sequence alignment.
Alpha and beta diversity analyses of the intestinal microbial community were performed using QIIME2. Alpha diversity measures the richness and evenness of microbial species within a sample.29 The Shannon and Simpson indices were selected to reflect community diversity, whereas the Observed_otus and Chao1 indices were used to evaluate species richness.30 For beta diversity analysis, the nonmetric multidimensional scaling (NMDS) method was adopted.31 Ordination plots were constructed using weighted and unweighted UniFrac distances to reveal differences in microbial community structure and their multidimensional distributions among samples.32–34
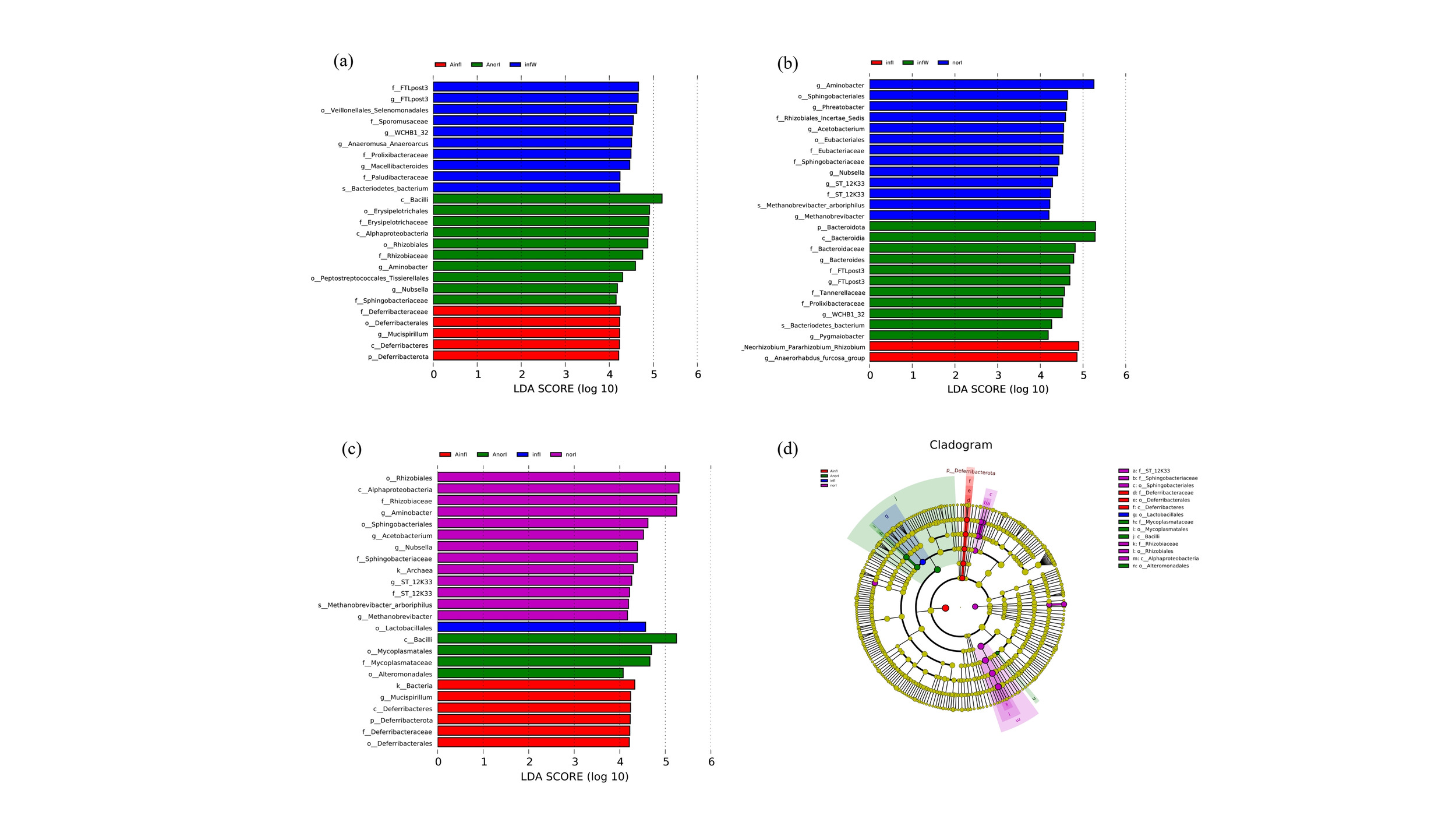
Linear discriminant analysis effect size (LEfSe) analysis was performed via LEfSe software (version 1.0),35 with the LDA score threshold set to 4 by default. The length of the bar plot represents the effect size of the differential species (i.e., LDA score). Functional prediction was performed using Tax4Fun software based on 16S rDNA sequencing data.36
2.5. Statistical analysis
The relevant charts were plotted using Origin 2021. The intestinal microbiota data were analyzed via the Novogene Magic Cloud Platform (https://magic.novogene.com/customer/main#/homeNew/3adfe99ed18ed30596999d5ce47b939a).
3. Results
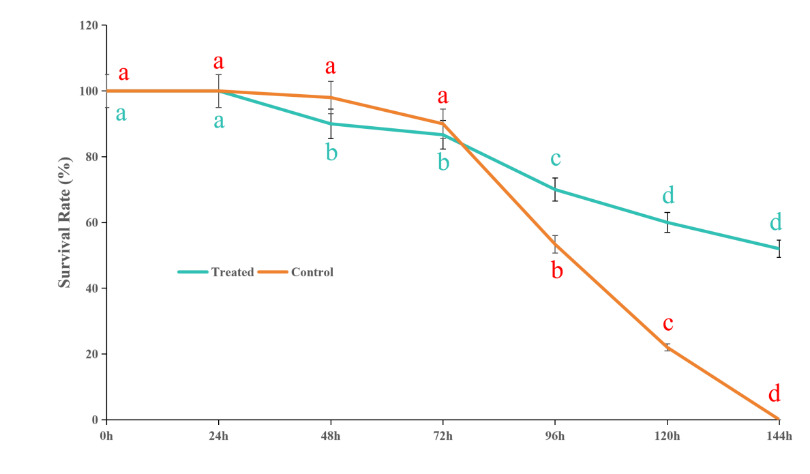
3.1. Effects of Astragalus polysaccharide on the survival rate of Whitmania pigra
During the experiment, individual deaths began to occur on the second day. By the sixth day, all individuals in the control group had died, while the survival rate of the W. pigra group injected with APS remained at 50%. The changes in survival rates across the experimental groups are shown in Fig. 1.
_and_the_c.png)
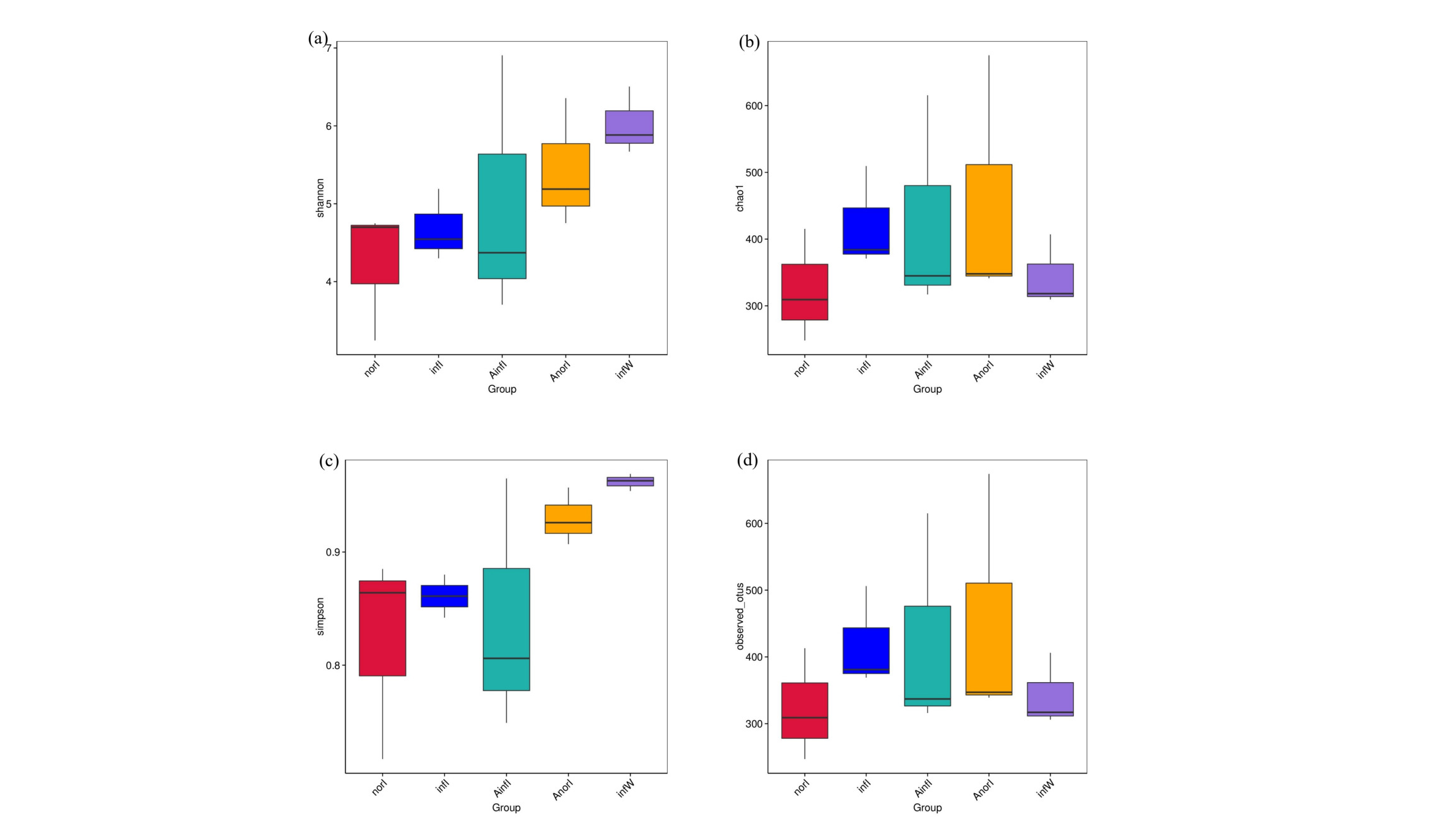
3.2. Effects of APS on the Alpha and Beta Diversity of the Intestinal Microbiota in W. pigra
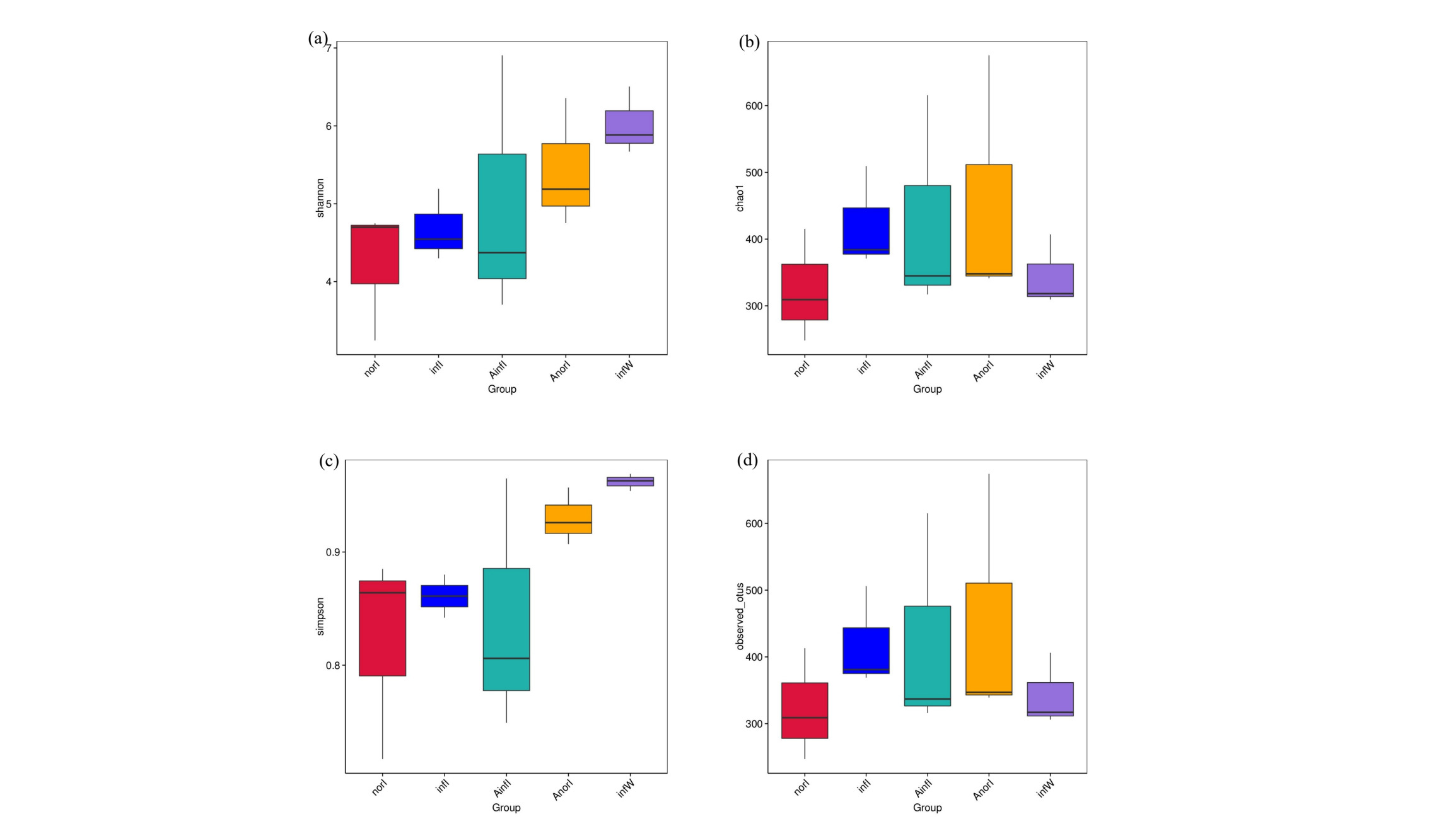
Detailed information on the diversity and richness indices of the intestinal microbiota in W. pigra is presented in Fig. 2. Compared with those in the control groups (infI and norI), the Shannon index (Fig. 2a), Chao1 index (Fig. 2b), Simpson index (Fig. 2c), and Observed_otus index (Fig. 2d), as well as the richness indices, were significantly greater in the treatment groups (AinfI and AnorI).
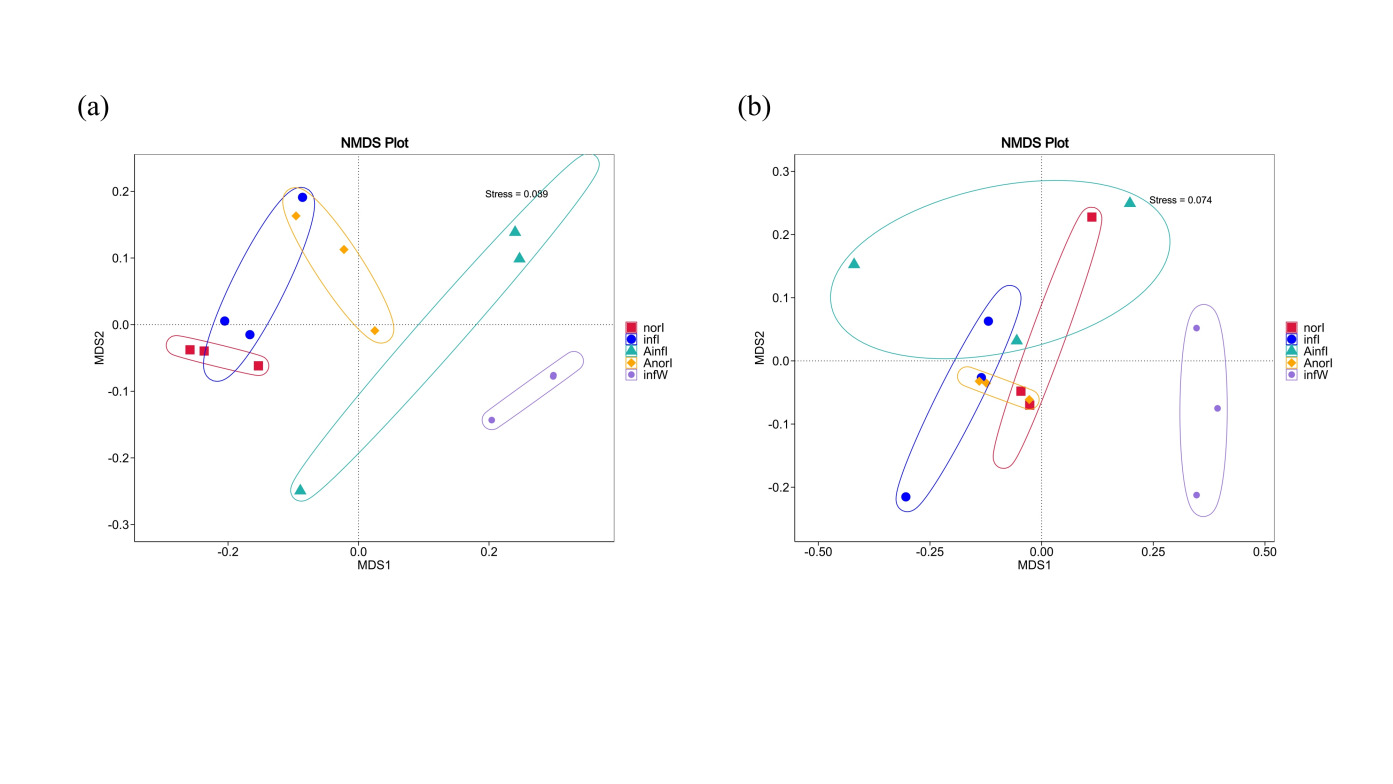
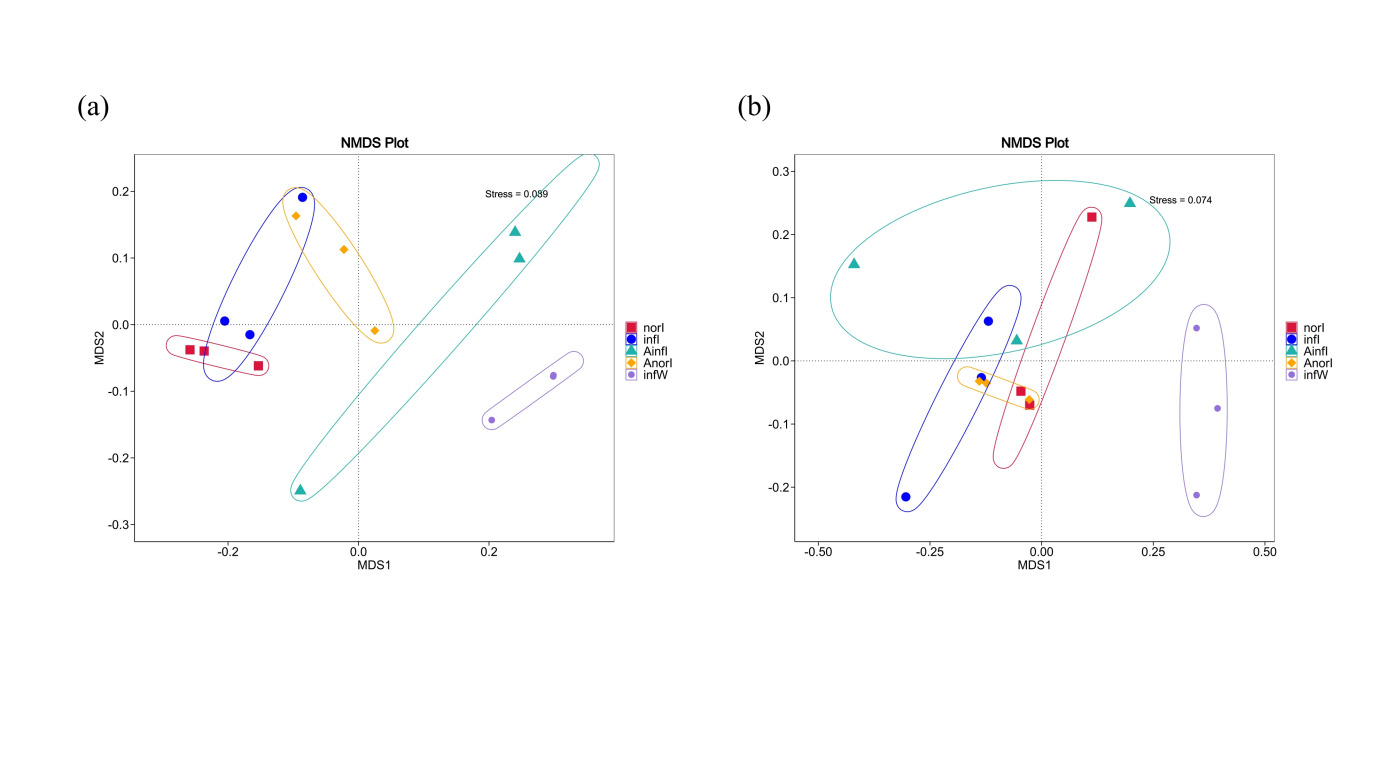
Beta diversity differences in the intestinal microbiota between the control and treatment groups of W. pigra were evaluated via nonmetric multidimensional scaling (NMDS) based on weighted UniFrac (Fig. 3a) and unweighted UniFrac (Fig. 3b) distances. The stress values of the NMDS plots were 0.089 and 0.074, respectively, indicating a good fit of the ordination results. The analysis revealed significant differences in microbial community structure between the two groups, with distinct clustering and separation in the plots.
3.3. Effects of APS on the Intestinal Microbiota Composition of W. pigra
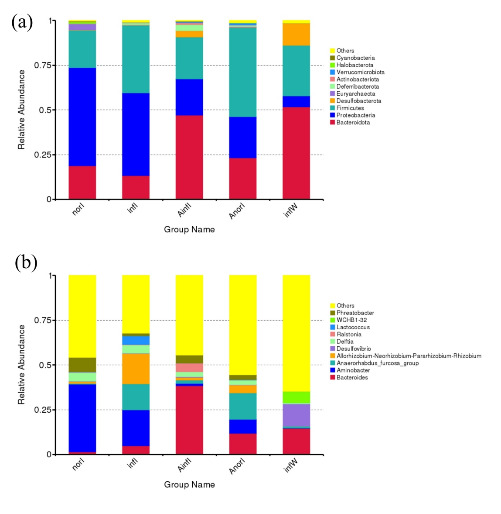
At the phylum level, the dominant phyla in the norI group were Proteobacteria, Firmicutes, and Bacteroidota. The dominant phyla in the infI group were also Proteobacteria, Firmicutes, and Bacteroidota. In the AinfI group, the dominant phyla were Bacteroidota, Firmicutes, and Proteobacteria. The dominant phyla in the AnorI group were Firmicutes, Bacteroidota, and Proteobacteria (Fig. 4a).
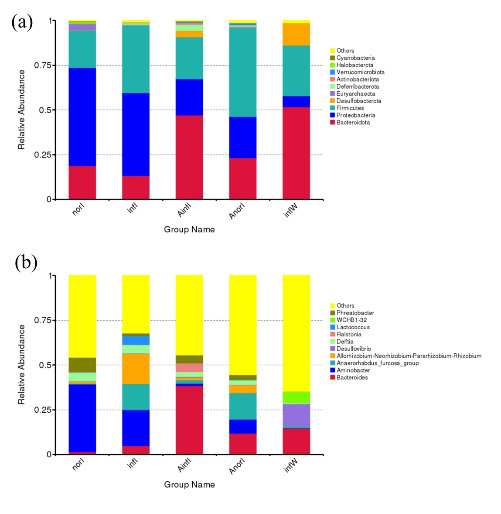
At the genus level, the dominant genera in the norI group were Aminobacter, Phreatobacter, and Delftia; those in the infI group were Aminobacter, Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium, and Anaerorhabdus_furcosa_group; those in the AinfI group were Bacteroides, Ralstonia, and Phreatobacter; and those in the AnorI group were Anaerorhabdus_furcosa_group, Bacteroides, and Aminobacter, respectively (Fig. 4b).
3.4. Differential abundance analysis of the intestinal microbiota composition in W. pigra
LEfSe analysis revealed that, at the genus level, Aminobacter and Nubsella were significantly enriched in the AnorI group compared with the other subgroups in the APS injection group (Fig. 5a), whereas Mucispillum was significantly enriched in the AinfI group. In the control group, comparisons within subgroups revealed that the Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium and Anaerorhabdus_furcosa_groups were significantly enriched in the infI group, whereas Aminobacter, Phreatobacter, Acetobacterium, Nubsella and Methanobrevibacter were significantly enriched in the norI group (Fig. 5b).
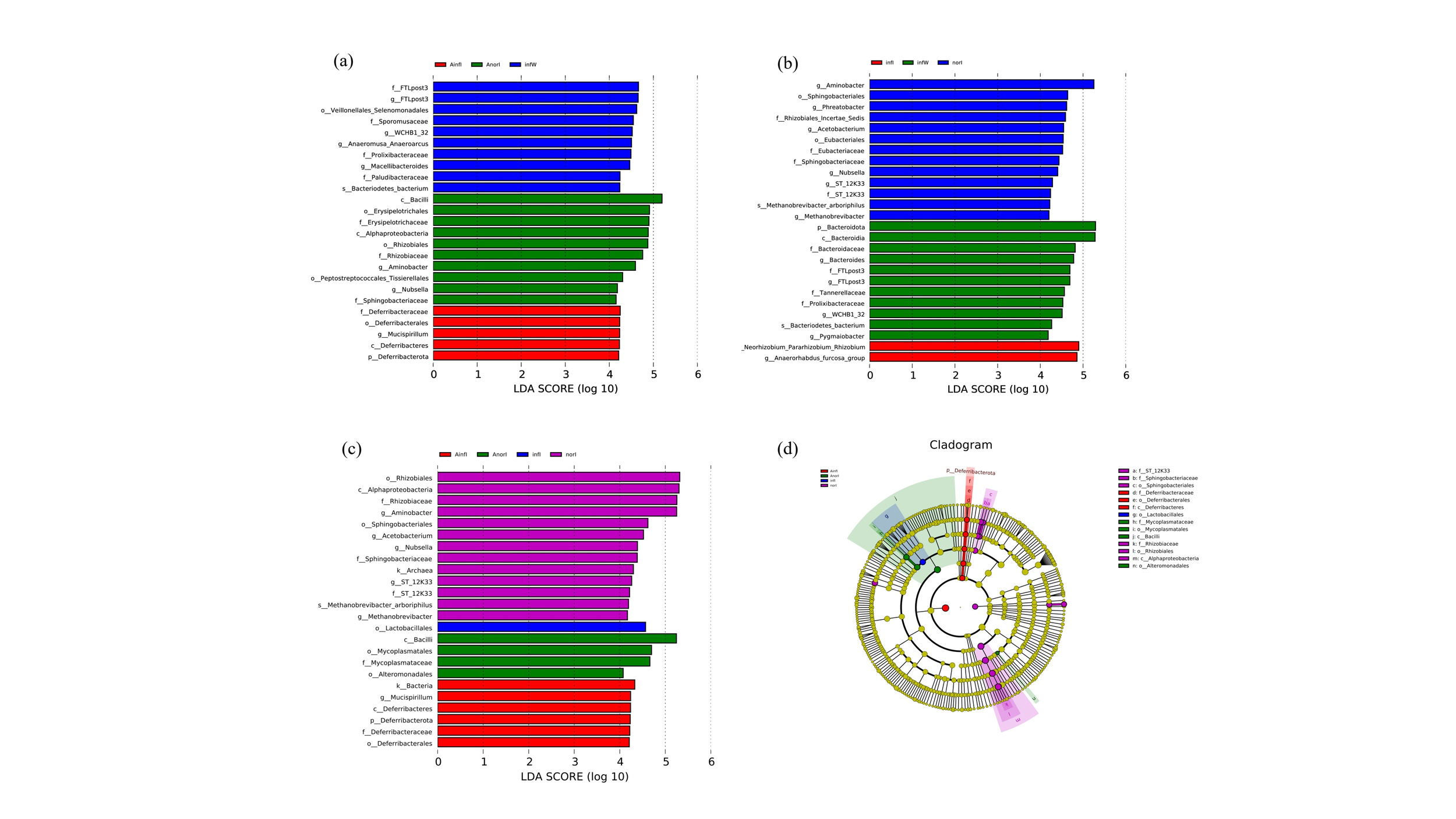
Pairwise comparisons among all four groups revealed that, at the genus level, Aminobacter, Acetobacterium, Nubsella and Methanobrevibacter in the norI group, as well as Mucispillum in the AinfI group, achieved relatively high LDA scores (Fig. 5c). Moreover, Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium in the infI group also had relatively high LDA scores (Fig. 5d).
3.5. Effects of APS on the intestinal microbiota of W. pigra
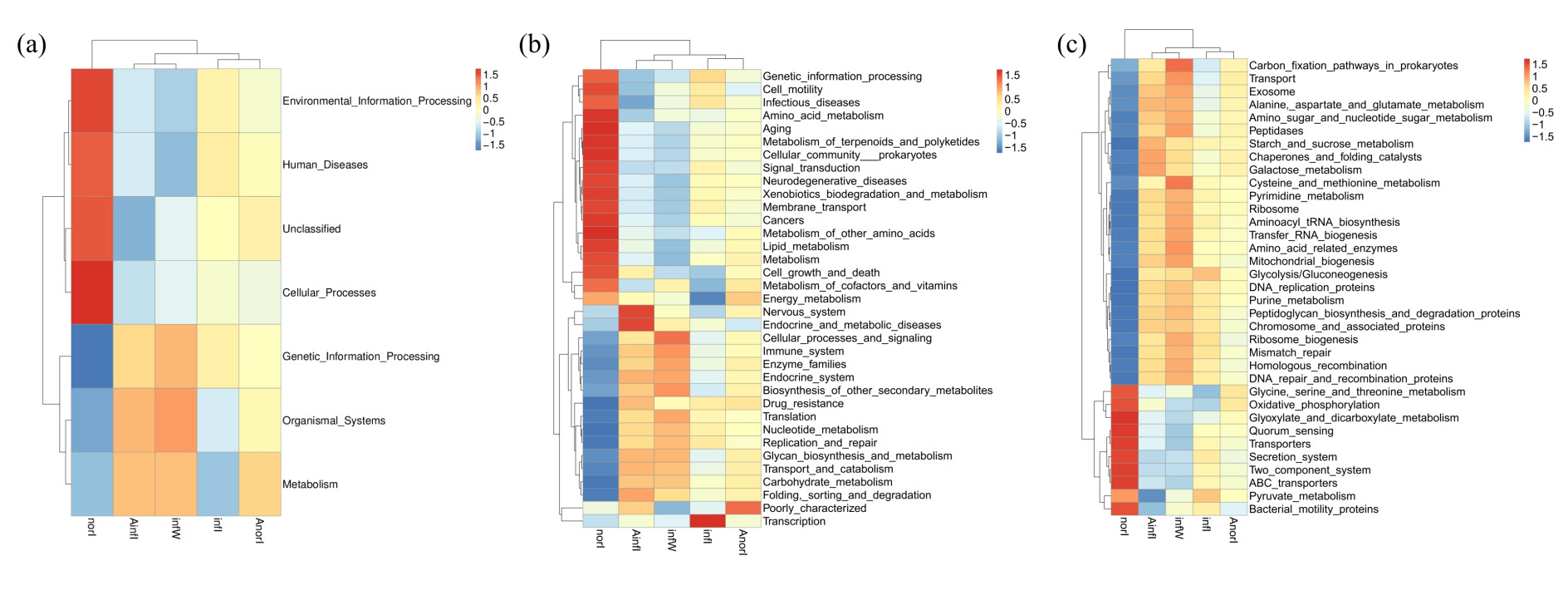
On the basis of the Tax4Fun tool and KEGG database, functional prediction and annotation of the metagenome represented by ASVs were performed. The functional pathways were classified into three levels (Level 1, Level 2, and Level 3) according to the KEGG hierarchical system. Among the intestinal samples from both the control and W. pigra treatment groups, metabolism was the most dominant level 1 functional category, followed by genetic information processing, environmental information processing, and cellular processes. Compared with the control group, the relative abundances of functional pathways related to metabolism and organismal systems in the treatment group were significantly higher, whereas those related to environmental information processing were significantly lower (Fig. 6a).
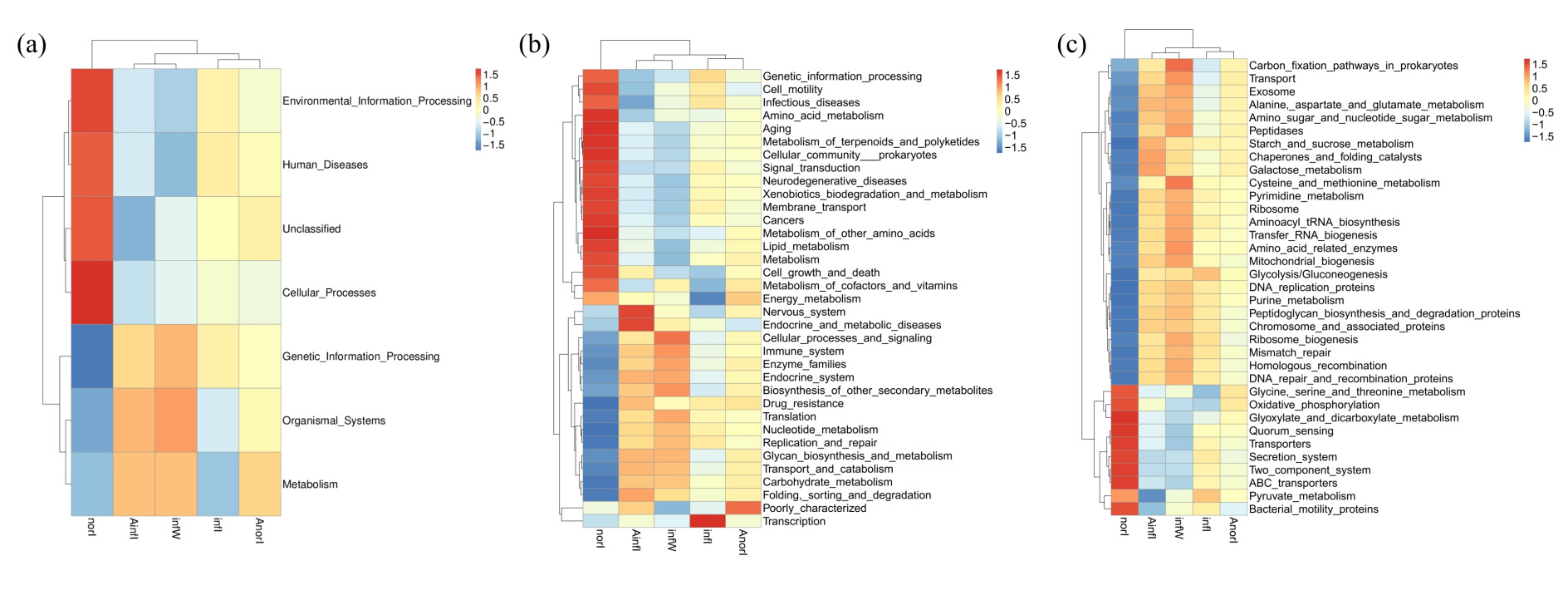
In the KEGG level 2 metabolic pathway classification, differences between intestinal samples before and after treatment were mainly reflected in pathways related to carbohydrate metabolism, membrane transport, and amino acid metabolism, whereas other gene functions were mainly involved in energy metabolism, nucleotide metabolism, and other pathways. The relative abundances of the above functional pathways differed significantly across all samples, indicating that APS treatment and disease progression significantly affected the functional composition of the intestinal microbiota in W. pigra (Fig. 6b).
In the classification of KEGG level 3 metabolic pathways of intestinal bacteria in the treatment group, the proportions of sequences related to carbon fixation pathways in prokaryotes; transport, amino sugar and nucleotide sugar metabolism; and galactose metabolism were significantly increased. In contrast, the abundances of pathways related to the secretion system and two-component system were significantly greater in the control group (Fig. 6c).
4. Discussion
Astragalus polysaccharide is a feed additive and immunoadjuvant widely used in aquaculture. Studies have demonstrated that it can not only enhance the immune efficacy of inactivated vaccines but also increase the activities of various immune-related enzymes and the expression levels of immune genes.37 When combined with inactivated vaccines, it can alleviate splenic tissue damage in rainbow trout (Oncorhynchus mykiss) infected with infectious hematopoietic necrosis virus (IHNV).38 A diverse microbial ecosystem is crucial for resisting pathogenic invasion.39 By improving the intestinal microbiota structure of aquatic animals, APS can increase the proportion of probiotics and the expression of intestinal immune factors, thereby enhancing disease resistance and survival rates. These effects have been verified in various aquatic species, including Pelteobagrus fulvidraco,40 Eriocheir sinensis and Acipenser dabryanus.17,41 In this study, we also found that APS treatment significantly increased the richness and diversity of the intestinal microbiota in W. pigra (Fig. 2). Furthermore, β-diversity analysis based on nonmetric multidimensional scaling (NMDS) revealed significant differences in community structure between the treatment group and the control group (Fig. 3) The intestinal microbiota of both the experimental group and the control group was predominantly composed of the phyla Bacteroidota, Proteobacteria, and Firmicutes, which is consistent with previous research.42 APS treatment altered the relative abundances of the dominant phyla: Bacteroidota and Firmicutes were the dominant phyla in the APS-treated group, whereas Proteobacteria was the most abundant phylum in the control group. Proteobacteria include many pathogenic species, and their high abundance is often associated with the development of intestinal diseases in the host. The decreased relative abundance of Proteobacteria and increased relative abundances of Bacteroidota and Firmicutes in the APS-treated group suggested that APS can reduce the colonization of pathogenic bacteria in the intestine, promote the proliferation of beneficial bacteria, and thus facilitate the recovery of intestinal health.
Compared with the control group, the treatment group showed a decrease in Proteobacteria relative abundance, whereas Bacteroidota relative abundance increased. Moreover, the abundance of Firmicutes in the AnorI group was significantly greater than that in the norI group. These results are consistent with the findings of previous studies on Ctenopharyngodon idellus,43 zebrafish, and Plectropomus leopardus supplemented with APS.44,45 At the genus level, the relative abundance of Bacteroides in the treatment group was significantly greater than that in the control group. This result aligns with the outcomes of studies involving Ctenopharyngodon idellus fed a Sudan grass diet and Pelophylax nigromaculatus fed a phytogenic diet.46,47 Bacteroides can decompose complex carbohydrates and produce SCFAs, which regulate the intestinal immune system,48 promote immune maturation, reduce inflammation,49 and inhibit pathogenic microorganisms, thereby improving intestinal health.50
The primary metabolic products of Bacteroidota are acetate and propionate, while Firmicutes mainly produce butyrate.51 The Bacteroides and Anaerorhabdus_furcosa_groups mentioned in this study belong to Bacteroidota and Firmicutes, respectively, and both are capable of producing SCFAs. The significant increase in the abundances of these two bacterial taxa can promote immune system maturation, inhibit proinflammatory pathways, and reduce the risk of intestinal inflammation. In addition, the abundances of Ralstonia, Mucispirillum and Aminobacter, which are members of Proteobacteria, distinctly changed in moribundly infected individuals: the abundances of the first two genera increased significantly, whereas that of Aminobacter decreased. Abnormal proliferation of Proteobacteria is widely recognized as a hallmark of intestinal dysbiosis.52 At present, the ecological functions of Aminobacter in invertebrate hosts remain unclear, and the underlying mechanisms for its downregulation during the disease course warrant further investigation. Bacteroidetes, Firmicutes and Proteobacteria play pivotal roles in host metabolism, and an imbalance in the ratio of these three phyla may induce metabolic disorders.53 Functional prediction analysis revealed that the intergroup differences were mainly concentrated in metabolism-related pathways, suggesting a close association between APS and the metabolic functions of the intestinal microbiota. These results demonstrate that APS can improve intestinal health and enhance host disease resistance in W. pigra by increasing the richness and diversity of the intestinal microbiota; regulating the relative abundances of dominant phyla, including Bacteroidota, Firmicutes and Proteobacteria; enriching beneficial bacterial genera that produce SCFAs; and inhibiting the proliferation of harmful bacteria.
Notably, intestinal microbiota analysis in this study was only performed at the experimental endpoint on day 6 post-challenge with Aeromonas allosaccharophila. At this time point, all W. pigra in the control group had succumbed to erysipelas, whilst the survival rate of individuals in the APS-treated group reached 50%. Although this sampling time point can highlight the greatest discrepancies in microbial communities between surviving and diseased leeches, sampling at a single time point makes it difficult to infer the temporal causal relationship between shifts in microbial composition and disease progression. It remains unclear whether the observed microbiota alterations occur before, concurrently with, or after the onset of erysipelas. Further research is recommended to adopt longitudinal gradient sampling on days 0, 2, 4, and 6 post-challenge to elucidate the dynamic succession of intestinal microbiota and clarify the intrinsic functional mechanisms linking APS intervention, intestinal microbial homeostasis, and disease resistance in W. pigra.
This study confirmed that APS injection effectively improved the survival rate of W. pigra infected with Aeromonas allosaccharophila. Comparative analysis of intestinal microbiota in leeches before and after infection revealed that astragalus polysaccharides optimized the intestinal microbial structure, increased the abundance of beneficial bacteria, and inhibited pathogenic bacteria. As a natural, safe, and effective feed additive and immunopotentiator, APS has great potential for preventing and controlling diseases in W. pigra aquaculture.
5. Conclusions
This study deepens understanding of the mechanism by which Astragalus polysaccharide regulates the intestinal microbiome of W. pigra and highlights the importance of research on the interactions between the active ingredients of traditional Chinese medicine and the microbiome. Although we have identified indicative microbial biomarkers, the specific molecular mechanism by which Astragalus polysaccharides affect host immunity and growth performance via the intestinal microbiota remains unclear. Furthermore, excluding the healthy blank control group (CK) from microbiota sequencing analyses impedes the reliable definition of the baseline microbiome of W. pigra, a notable limitation that ought to be addressed in future research. Future research should focus on the regulatory network among Astragalus polysaccharide, microbiota, and host, so as to improve the systematic understanding of the mechanism by which traditional Chinese medicine ingredients regulate the health of aquatic animals. In addition, although Astragalus polysaccharide shows application prospects as a potential prebiotic, dose-response experiments and long-term intervention studies are still needed to evaluate its optimal efficacy in regulating intestinal flora structure and enhancing disease resistance. Given the urgent need to reduce antibiotic use in aquatic animal farming, an integrated health management strategy combining traditional Chinese medicine with modern microbiome technologies is of great significance for the sustainable development of aquatic economic species such as W. pigra.
Acknowledgement
This research was financially supported by the National Natural Science Foundation of China (NSFC, grant no. 82474041 and 82073968), China; The Doctoral Program of the Science Research Foundation of Guizhou Education University (2024BS006 and 2024BS011), China; Rewardand subsidy fund project of Guizhou Education University, Ministry of science and technology of the people’s Republic of China and National Natural Science Foundation of China (2023GZJB004), China. Guizhou Forestry Administration Scientific Research Project (QLKH [2023] 11 and QLKH [2025] 11), China; Guizhou Science and Technology Support Plan Project ([2023]188), China.
CRediT authorship contribution statement
Conceptualization: Kai Ma. Methodology: Kai Ma. Data curation: Fei Liu. Formal Analysis: Bo Huang, Mengxue Li, Chao Feng, Wenping Yang. Investigation: Guiyan Shao, Hongwei Ao. Software: Kai Ma. Writing – original draft: Kai Ma. Resources: Fei Liu. Validation: Chenbo Sun, Hong-Yan Tian. Writing – review & editing: Boxing Cheng. Project administration: Boxing Cheng. Supervision: Kui Zhang.
Ethics approval
Ethical approval is not required for the paper.
Declaration of competing interest
The authors declare no competing interests.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.