1. Introduction
Fish oil has been the main lipid source used in the formulation of many carnivorous fish and crustaceans feed, owing to its ready availability, unique high palatability and rich in n-3 long-chain polyunsaturated fatty acids (n-3 LC PUFAs), such as eicosapentaenoic acid (EPA, 20:5n-3) and docosahexaenoic acid (DHA, 22:6n-3).1–3 These fatty acids have been proven to be essential for the formation of new cellular structures and for survival, growth, morphogenesis, predator behavior, pigmentary processes, and bone formation in fish larvae.4,5
With the global expansion of marine finfish and crustacean aquaculture sectors, the demand for fish oil in aquaculture is predicted to increase by about 14%, an estimated 908,000 tons.6,7 However, the total annual production of fish oil from forage fish has remained stable about 1~1.25 million tons, of which 75% was used in aquafeeds and is not expected to increase in the future due to dwindling catches of capture fisheries.7 As such, the global fish oil supply is one of the limiting factors for the aquaculture feed industry. Thus, the efforts to replace fish oil with alternative lipid sources are required to promote sustainable expansion of the aquaculture industry.7,8 The replacement of fish oil with vegetable oils (such as soybean oil, linseed oil, rapeseed oil and palm oil) has been extensively researched, but high levels of these oils may cause unwanted secondary effects in fish health and flesh quality, due to the lack of n-3 PUFAs.2,9–12
Microalgae are important primary producers in aquatic ecosystems and play a vital role in maintaining the entire food chain.13 It has been shown that microalgae are rich in crude protein (21%–71% dry weight), lipid, n-3 PUFAs, carbohydrates (10%–25% dry weight), vitamins and minerals.14 Moreover, it had many bioactive molecules, such as polysaccharides, β-carotene, astaxanthin, lutein, neoxanthin, and zeaxanthin, which bring a more positive impact on growth, reproduction, immune response, disease resistance, and pigmentation of aquatic animals.13,14 Based on its better nutritional and medicinal properties, microalgae had been considered as a potential protein resources and functional additives in animal feed.15 Thraustochytrids, such as Schizochytrium, are eukaryotic and heterotrophic microorganisms found widely in marine and other saline environments, and accumulate triacylglycerols rich in DHA and EPA in their cells.9,16 Marine thraustochytrid strain has been shown to be effective in enriching Artemia nauplii and rotifers prior to feeding them to fish larvae.16,17 Large-scale production of heterotrophic microalgae by established fermentation technology is now commercially available, such as Schizochytrium sp, with high lipid content (55%-75% dry matter) and up to 49% DHA of total lipids and high percentage of n-3 LC-PUFA rich lipid, which is used as an alternative source to fish oil for n-3 LC-PUFAs and fish meal in aquatic feeds.9–11,15,18–23
Rainbow trout is a carnivorous cold-water fish that has been distributed worldwide and extensively cultivated in China because of its better flesh quality, such as high protein, low fat and rich in DHA, EPA.24 Microalgae play an important role in the rearing of aquatic animals and serve as the main nutrition source for the larval stages of mollusks, shrimp, and fish.13,25 Ingestion rates of microdiets are often lower than those of live prey during the first days of feeding, which may limit the availability of nutrients for normal growth and development. This can be interpreted as pancreatic and intestinal enzyme secretion not being sufficiently stimulated, and the incomplete development of the digestive tract of fish larvae. Data pertaining to digestive enzyme activity of aquatic animals may help to better explain nutrient use and relative digestibility, and help overcome nutritional problems associated with the formulation of diets that best meet an animal’s nutritive capability.26–28 However, there is no relevant report on the potential of Schizochytrium meal as a source of DHA in rainbow trout larvae microdiets. Therefore, this study was designed to evaluate the effects of replacement of fish oil with DHA-rich Schizochytrium meal on growth performance, digestive enzyme activity, body fatty acid composition and intestinal histology of rainbow trout larvae.
2. Materials and methods
2.1. Diet formulation and preparation
Schizochytrium meal was obtained from Zhihe Biotechnology Co., Ltd. (Changzhou, Jiangsu, China), and it contained 13.85% crude protein and 39.83% crude lipid. In order to maintain the need of fish growth, dietary DHA level was fixed at 0.8% dry matter.29 Five isonitrogenous (crude protein, 52%) and isolipidic (crude fat, 15%) diets were formulated, in which 0% (R0), 25% (R25), 50% (R50), 75% (R75) or 100% (R100) of fish oil DHA was replaced by Schizochytrium meal (Table 1). Soybean oil was added as dietary lipid source to make all the diets isolipidic. Composition and nutrient levels of experimental microdiets are shown in Table 1. The fatty acid compositions of Schizochytrium meal and test diets are given in Table 2.
Microdiets were manufactured by micro-bonding technology. Ingredients were ground to a fine powder that passed through a 96 μm screen. Then the ground ingredients were blended thoroughly and stirred with oil and water to produce stiff dough. Diets were pelleted by passing the dough through an experimental feed mill and dried for about 12 h in a ventilated oven at 60°C. After drying, the diets were broken up, sieved into proper pellet sizes (150~250 μm and 250~380 μm), and then sealed in plastic bags and stored at −20°C until used.
2.2. Feeding trial
The rainbow trout larvae (initial body weight: 0.25 ± 0.01 g) were obtained from a commercial hatchery (Weifang, China) and reared at the Aquaculture Research Center, Rizhao Polytechnic (Shandong, China). Fish larvae were randomly allotted to 15 tanks (100 × 45× 80 cm) with 500 fish to per tank. Each test diet was randomly assigned to triplicate tanks. Fish were hand fed to apparent satiation four times daily at 6:00, 10:00, 14:00 and 18:00, respectively. During the 56-day experimental period, temperature was 14 ± 0.5°C, dissolved oxygen level above 6.5 mg L−1, pH was 6.8~7.2, ammonia-N was less than 0.1 mg L-1, the photoperiod was set at 12-h light and 12-h dark. Mortality and feeding behavior were monitored every day.
2.3. Sample collection and analysis
At the end of the experiment, all fish were fasted for 24 h before harvest. The total number of fish in each tank was measured to determine the survival. 15 fish were randomly sampled from each tank to measure final body weight (FBW) and final body length (FBL), respectively. Fifty fish were randomly collected from each tank, and then immediately stored at −80°C for enzyme activity assays. 20 fish in each tank were dissected, mixed and stored in a plastic bag, and then stored at −20°C for fatty acid composition analysis. Intestines of three fish each tank were sampled and stored in 4% paraformaldehyde fix solution for H&E analysis.
The fish were dissected to separate pancreatic and intestinal segments on a glass plate maintained at 0°C. For enzymatic assays, the dissected samples were homogenized in 2 ml cold ultrapure water (0°C). The homogenates were centrifuged at 4°C at 3300 ×g for 10 min, and then the supernatant was gently collected and frozen at −80°C until analysis of enzyme activity. Purified brush border membranes (BBM) from the homogenate of the intestinal segment were obtained according to a method described by Crane et al.30 Before CaCl2 solution addition, 1 mL of the homogenate was diverted for intestinal enzyme assays. After addition of 0.1 M CaCl2, the homogenate was centrifuged at 3300 ×g for 10 min in a centrifuge at 4°C. The supernatants were kept in new vials and stored frozen at −80°C until assayed for the digestive enzymes activities or protein content.
The trypsin, amylase, lipase, leucine-aminopeptidase (LAP) and alkaline-phosphatase (AKP) activities were analysed using reagent kits (Jiancheng Bioengineering Institute, Nanjing, China) according to the product manual. Protein concentration was determined by the Bradford procedure using bovine serum albumin (BSA, A-2153, Sigma) as a standard. All enzyme activities were carried out in triplicate, and expressed as specific activity (U mg/protein). The fatty acid profiles were analyzed using the detail procedures described by Zuo et al.31 Briefly, about 100 mg freeze-dried muscle samples were added into a 10 ml volumetric screwed glass tube with plastic cover. Then 3 ml potassium hydroxide methanol (1 N) was added and heated on 75°C water bath for 25 min. After that, 3 ml HCL-methanol (2 N) was added, and the mixture was heated on 75°C water bath for another 25 min. At last, 1 ml hexane was added into the mixture above, shaken vigorously for 1 min, and then allowed to separate into two layers. Fatty acid methyl esters in the upper layer were separated, and quantified by HP6890 gas chromatograph (Agilent Technologies Inc., Santa Clara, California, USA) with a fused silica capillary column (007-CW, Hewlett Packard, Palo Alto, CA, USA) and a flame ionization detector. The column temperature was programmed to rise from 150°C up to 200°C at a rate of 15°C min-1, and from 200°C to 250°C at a rate of 2°C min-1. Injector and detector temperature were 250°C, respectively.
H&E staining was used to analyze the change of intestinal structure and liver histology of fish fed a diet with Schizochytrium meal. The fixed intestinal and hepatic samples were sheared, dehydrated in a series of ethanol solutions, infiltrated in xylene and embedded in paraffin, sliced into 4 μm thick sections, stained with H&E. The sections were scanned using a 3D panoramic scanner (Pannoramic, 3Dhistech, Hungary, Japan). The morphological structure of intestine and liver was imaged with a Pannoramic digital pathology system (CaseViewer2.4, 3Dhistech, Hungary) and measured using an analysis software (Image-Pro Plus 6.0, Media Cybemetics, USA).
2.4. Calculations and statistical analysis
Survival (%) = final fish number/initial fish number
Weight gain (WG, %) = (Wf − Wi) ×100/ Wi
Specific growth rate (SGR, % day-1) = (Ln Wf − Ln Wi) ×100/t
Where Wf is final body weight (g), Wi is initial body weight (g), t is experimental duration (d).
SPSS for Windows software version 21.0 (SPSS Inc., Chicago, Illinois, USA) was used to analyze all data. If a significant difference between the treatments was detected, it was subjected to one-way analysis of variance (one-way ANOVA). When significant differences were found (P<0.05), Duncan’s multiple range test was used to compare the mean values between individual treatment. The data are presented as means ± S.E (n = 3).
3. Results
3.1. Growth performance
After the 56-day feeding trial, the overall survival was 74.13% to 76.60% and was not affected by dietary treatments (P>0.05). Fish fed Schizochytrium meal-added diets (R25, R50, R75 or R100) had similar final body length, WG and SGR to fish fed control diet R0 (P>0.05). These results showed that Schizochytrium meal could replace 100% fish oil DHA in the diet without negatively affecting growth (Table 3).
3.2. Digestive enzyme activity
Trypsin, lipase and amylase activities in pancreatic and intestinal segments were not affected by dietary treatments (P>0.05). Also, no significant differences were found in AKP and LAP activity in intestinal and in purified brush border membrane of intestine among dietary treatments (P>0.05) (Table 4).
3.3. Fatty acid composition
The percentages of all the identified fatty acids in fish fed diets are presented in Table 5. There were no significant differences in C16:0, MUFA, SFA, DHA and n-3 PUFAs content in fish samples among dietary treatments (P>0.05). C14:0, C16:1n-7, EPA, C20:4n-6 content, n-3/n-6 ratio significantly decreased, while C18:0, C20:0, C18:1n-9 content, DHA/EPA ratio significantly increased in fish as dietary algae meal level increased from 0% to 100% (P<0.05). C18:3n-3, C18:2n-6, PUFA and n-6 PUFAs in fish increased with increasing algae meal replacement level from 0 to 100% (P<0.05).
3.4. Intestinal histology
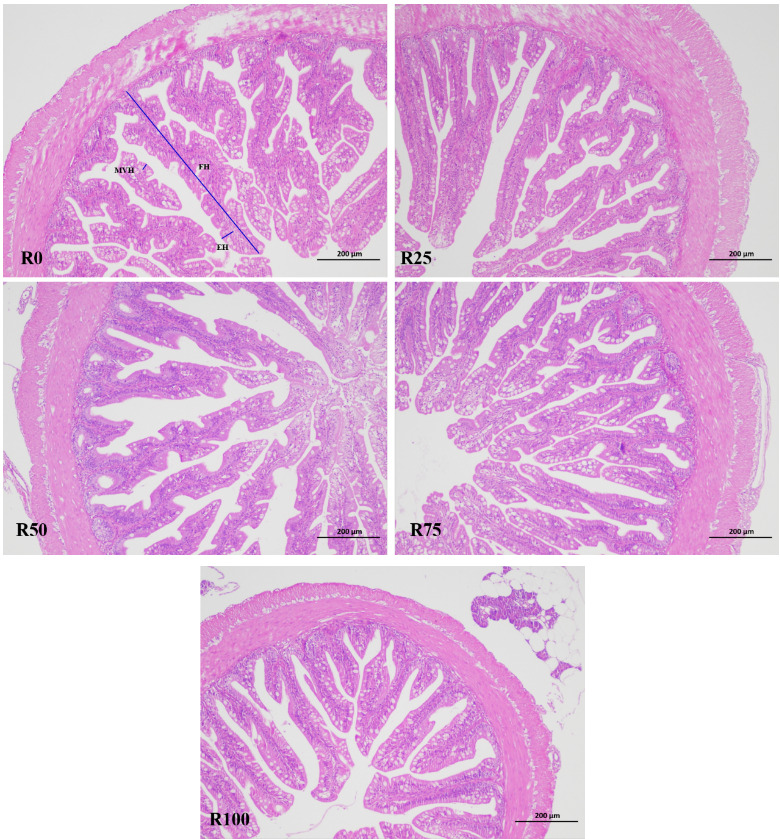
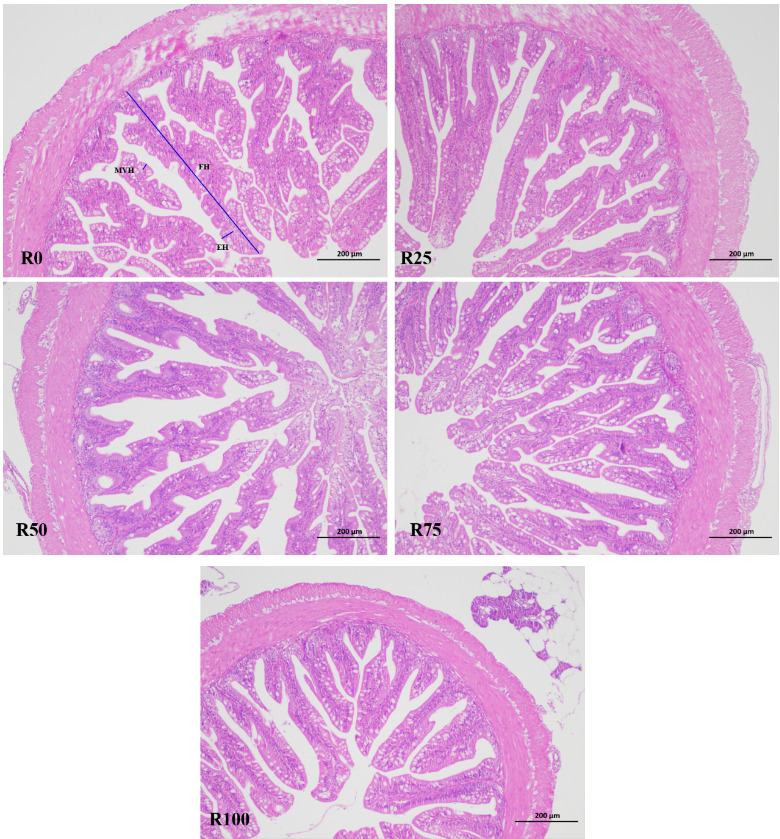
As shown in Table 6 and Figure 1, there were no statistically significant differences in intestinal fold height, enterocyte height and microvillus height among all dietary treatments (P>0.05).
4. Discussion
4.1. Growth performance
Single cell oils, such as oil from thraustochytrids, diatoms, and other groups, can produce high concentrations of n-3 LC-PUFA, showing great potential to provide a renewable source of these fatty acids.10 This study showed that 100% of dietary fish oil DHA could be replaced with Schizochytrium meal in microdiets of rainbow trout larvae without detrimental effects on survival and growth performance, and this demonstrated Schizochytrium meal as an alternative DHA source for fish oil in microdiets of rainbow trout larvae. Similar result have been reported by Carvalho et al. who found that oils from microalgae rich in n-3 LC-PUFA could 100% replacement of fish oil in combination with poultry oil and rapeseed oil for juveniles sea bream.1 Zatti et al. also found that diet with 11.2% Schizochytrium sp. oil could 100% replace fish oil without negative effects on feed intake, growth and feed utilization of Atlantic salmon grower.32 Similar results have also been reported in Atlantic salmon (Salmo salar),10,18 gilthead seabream (Sparus aurata),11,33 Pacific white shrimp (Litopenaeus vannamei)21 and olive flounder (Paralichthys olivaceus).34 Moreover, Neylan et al. reported that growth increased with increasing Schizochytrium sp. level in fish meal-free diets for sablefish (Anoplopoma fimbria), and fish fed diet with 12% microalga had similar performance to fish fed fish meal diet.35 Similarly, studies on gilthead seabream19,36 and white shrimp37 also observed that fish oil replacement by n-3 PUFAs rich microalgae led to higher survival, growth and stress resistance. This could be interpreted as the presence of biologically active molecules, such as pigments (carotenoids and phycobilins), polysaccharides, proteins, peptides, nucleic acids, fatty acids and phospholipids in microalgae.14 On the contrary, studies on gilthead seabream larvae11,19 and Atlantic salmon20,38 found that fish oil substitution by algal meal in diets exhibited lower growth and survival, and this could be related to an epithelial degeneration observed in the anterior intestine, and this degeneration could be related to the disorders of lipid metabolism and the higher lipid content in fish.19 In addition, the imbalance of fatty acids and a lower digestibility may impair growth and development of aquatic animals.39,40 The addition of EPA sources to diets with total replacement of fish oil by microorganism produced DHA seems to be necessary to further promote larval growth performance.11,33 Furthermore, microalgae possess a very rigid cell wall, resulting in a low bioavailability of intracellular components.41 Nevertheless, Teuling et al. found that the nutrient digestibility of the unicellular sources (microalgae and cyanobacteria) did not relate to the mechanical cell wall hardness.42 The application of algal meal to replace fish oil in diets for aquatic animals has provided variable results. These disagreements may be related to differences in animal species, life stage, nutrition digestibility, microalgae meal species, fatty acids composition and culture conditions.
4.2. Digestive enzyme activity
Changes in digestive enzyme profile have been used as indicators of the effects of dietary composition on the onset of maturation processes in the digestive tract.43 In this study, no significant differences were observed in trypsin, lipase and amylase activities in pancreatic and intestinal segments among fish fed different diets. This indicated that the replacement of fish oil with Schizochytrium meal may not significantly affect enzyme secretion of rainbow trout larvae. However, Vizcaíno et al. found that gilthead sea bream fed 12% microalgae meals showed higher trypsin activity than those fed on a microalgae-free diet.44 Lipase activity in the hepatopancreas of white shrimp increased with increasing dietary Schizochytrium sp. level from 28.8 to 88.5 g/kg, whereas adipocytes decreased.21 Algae growth regulators, such as polyamides, have been shown to stimulate cholecystokinin release in rats, which mediates the release of pancreatic enzymes.45
Brush border membrane (BBM) enzyme assays have been successfully used to determine the degree of the maturation process of the digestive function in the intestine of fish larvae.46,47 LAP and AKP are regarded as indicators for a well-differentiated intestinal BBM and have previously been found to exhibit high activities in fish larvae fed appropriate diets.47,48 In this study, specific activities of AKP and LAP in the intestinal segment and purified BBM of the intestine were not affected by dietary treatments. However, Vizcaíno et al44 found that AKP and LAP activities increase in fish fed diets with Scenedesmus almeriensis, and this may be related to maturation of the intestinal membrane and enhanced survival in fish.46 Data comparisons on digestive enzyme activities between studies can be difficult due to the variability in collection and analytical methods as well as other factors such as fish size, environment and nutritional status.49
4.3 Fatty acid composition
A number of trials have demonstrated that the fish fillet fatty acid profiles were influenced by diet treatment, generally reflecting the fatty acid profiles of the diet.35,38 Many studies using plant-based diets have shown a decrease in the levels of flesh EPA and DHA, due to the lack of these fatty acids in plant-based ingredients.8,34,37,38 In this study, no significant differences in body DHA content between fish fed a control diet and diets with Schizochytrium meal replaced 25%-100% of fish oil, which indicated that n-3 PUFAs in Schizochytrium meal were well digested and absorbed as an alternative source of DHA. Previous studies on channel catfish (Ictalurus punctatus),8 Atlantic salmon,10 sablefish,35 and gilthead seabream1,19,33,36 denoting that dietary fish oil replaced with thraustochytrid oil significantly increased fillet DHA levels. In this study, EPA content significantly decreased as dietary microalgae meal increased from 0% to 100%. Similar results were observed in Atlantic salmon,10,18,20,32,38 seabream,11 and gilthead seabream,33 which may be due to the absence of EPA in algae-based diets. However, some authors reported that EPA levels in fish were not affected by diets containing algae meal.19,34,35 The hepatopancreatic linoleic acid, linolenic acid and DHA levels of white shrimp increased with increasing dietary Schizochytrium sp. level, while EPA levels decreased.21
The most common problem with substituting fish oil with available alternatives is the unavoidable modification of fillet fatty acid composition, and in especial leads towards a reduction in n-3 LC-PUFA an increase in n-6 PUFA content.50 Research has shown that feeding the dried algae increases n-3 LC-PUFAs in the breast meat of broiler chickens and in the milk of dairy cows.51 Similar to the studies from Atalah et al,19 Kousoulaki et al,20 Zatti et al.,32 Qiao et al,34 and Neylan et al.,35 we also found n-6 PUFAs increased, while no significant differences in n-3 PUFAs were observed when rainbow trout were fed algae-based diets. Therefore, replacement of fish oil with Schizochytrium meal maintains the EPA and DHA content of fish, both of which are beneficial to human health.
Given that many plant-based feed ingredients contain a low n-3/n-6 ratio, the challenge is to promote good animal performance, and maintain a high n-3/n-6 ratio, which plays a positive role in human health and is also important in aquafeeds.2,52 In this study, the n-3/n-6 ratio in fish decreased with increasing algae meal replacement level. Similarly, in comparison with fish oil fed fish, n-3/n-6 ratio was lower in Schizochytrium-fed larvae.11,32 However, Qiao et al reported that the substitution of fish oil with microalgae increased n-3/n-6 ratio in olive flounder.34 Atalah et al and Neylan et al. observed that seabream postlarvae fed diet with algae did not affect the n-3/n-6 ratio.19,35
4.4. Intestinal histology
The intestinal tract is the main site of nutrient digestion and absorption, hormone secretion, and immune protection in fish, and the intestinal structure varies with the diet.53 The appropriate intake of microalgae can increase the intestinal folds and villus number, villus height of fish and shrimp, promote the development of the intestines, and this might be related to the active substances, such as spermine. Spermine can be rapidly absorbed by the intestinal mucosal epithelial cells, promote the proliferation of intestinal cells, and had positive effects on the intestinal development of fish and may promote the digestion and absorption of nutrients.54,55 Microdiet with 0.10% spermine could increase intestinal microvilli length and mucosal layer thickness of half-smooth tongue sole (Cynoglossus semilaevis) postlarvae.56 In this study, no clear differences were noted on intestinal fold height, enterocyte height and microvillus height among fish fed different diets. Similarly, a diet containing 10% pre-extruded microalga has no effects on histomorphology of distal intestine of Atlantic salmon, but increased the cell proliferation in intestinal villi.57 Neylan et al. found that diets with up to 12% Schizochytrium sp. were well tolerated for sablefish and did not cause histomorphological changes in the distal intestine.35 Kousoulaki et al. also support similar findings with no negative impacts found in the hind intestine histology of salmon.20,58 Bou et al. showed that high DHA supplementation had a stronger ability to support normal intestinal structures and function than EPA.59 These results indicated that microalga meal inclusion diets were tolerated and consistent with healthy intestinal. Further studies should focus on immunohistochemistry to gain a deeper understanding of tissue homeostasis.
5. Conclusion
In conclusion, the current study showed that 100% fish oil can be replaced by Schizochytrium meal without affecting survival, growth performance, digestive enzyme activities, and health of rainbow trout larvae. Including Schizochytrium meal did not affect the amount of DHA and n-3 PUFAs in fish, and Schizochytrium meal could be used as a viable DHA source for alternative rainbow trout microdiets.
Acknowledgments
This study was financially supported by a grant from the Innovative Team Projects of Scientific Research in Rizhao Polytechnic (2025KC03).
Authors’ Contribution - CRediT
Writing – original draft: Bin Hu (Lead). Methodology: Ning Fu (Equal), Shengwen Niu (Equal). Project administration: Ning Fu (Lead). Formal Analysis: Jing Jia (Equal), Meiling An (Equal), Mengxin Xing (Equal). Investigation: Jing Jia (Equal), Mengxin Xing (Equal). Resources: Lu Zhao (Lead). Writing – review & editing: Yuyu Wang (Lead). Funding acquisition: Yuyu Wang (Lead). Supervision: Yuyu Wang (Lead).
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
All experimental protocols were approved by the Animal Care and Use Committee of Rizhao Polytechnic and followed the laboratory animal guidelines for ethical review of animal welfare in China (GB/T 35892- 2018).
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.