Introduction
The microbiome is a critical determinant of animal health, mediating essential biological functions such as energy metabolism, immune development, growth regulation, nutritional processing, and stress responses.1,2 Evidence suggests that the intestinal microbiota can be affected by developmental stage and gender, diet, habitat, salinity, etc.3–7 As global temperatures rise, increasing attention is being directed toward understanding how temperature influences the gut microbiota of aquatic organisms. For example, Greenspan et al.8 reported that warming shifted environmental bacteria and community composition, which in turn promoted dysbiosis and stunted tadpole growth. Zhu et al.9 indicated that elevated environmental temperatures may have indirect impacts on both the alpha and beta diversity of the salamander microbiome. In aquatic invertebrates, Li et al.10 showed that thermal stress (27℃) altered the gut microbiome of Mytilus galloprovincialis, increasing opportunistic pathogens (Vibrio, Arcobacter) and mortality. Similarly, in sea cucumbers Apostichopus japonicus, Zhao et al.11 found seasonal warming-induced aestivation shifted gut microbiota: reduced dietary glycan-preferring taxa, increased mucin-degrading bacteria and opportunistic pathogens, with intestinal atrophy and enhanced immunity. Few existing studies have also revealed that temperature can alter the flavor and ecological adaptation of invertebrates by modulating intestinal microbiota.10,12 Nevertheless, studies aimed at investigating the relationship between gut microbiota and temperature in aquatic invertebrates are still limited. Therefore, there is an urgent need to explore further the effects of temperature on the gut microbiota of different species of invertebrates, particularly those that are economically important.
The swimming crab Portunus trituberculatus is an important economic crab that commonly inhabits reefs and gravels in the offshore waters of Japan, Korea and China.13 As a commercially significant crustacean, P. trituberculatus is renowned for its high nutritional value. In 2022, its aquaculture production reached approximately 109,017 metric tonnes, making it one of the most crucial cultured crustaceans in China.14,15 As an ectotherm, P. trituberculatus is highly sensitive to temperature and occupies a significant ecological niche. Previous studies showed that appropriate temperatures can stimulate metabolic physiology, ovarian development, molting, and nonspecific immunity of P. trituberculatus.16–18 Moreover, the intestine serves as a vital organ with significant physiological functions in P. trituberculatus.19 Nevertheless, the effects of temperature on the intestinal microbiota of P. trituberculatus remain poorly understood. To address this gap, we aimed to clarify how temperature fluctuations regulate the intestinal histological structure and microbial community of P. trituberculatus and further explore their links to metabolic function. The results will not only address a critical gap in our understanding of this species’ aquaculture health but also provide valuable insights into the adaptive responses of invertebrate gut microbiomes to thermal stress, a phenomenon of increasing relevance in the context of climate change.
To investigate the physiological responses of P. trituberculatus during acclimation to seawater temperature, we analyzed the effects of temperature stress (20℃, 23℃, 26℃, 29℃) on the intestinal histology and microbiota of P. trituberculatus using Illumina HiSeq sequencing of the 16S rRNA gene. The findings of this study will provide insights into the diversity and distribution of intestinal microbes and help explore the potential relationships among metabolic function changes, intestinal microbes, and temperature.
Materials and methods
Experimental crabs
The experimental crabs were obtained from the outer pond of the research base of Zhejiang Fisheries Research Institute, Zhejiang, China. Female crabs (body weight: 135 ± 25 g), which had finished the puberty molt, were selected for the experiment. Prior to the experiment, all crabs were visually inspected for signs of disease or other health issues, and only those appearing healthy and free of lesions or abnormal behavior were selected. During the experiment, each crab was individually reared in plastic water tanks (Length×Width×Depth=1.84 m×1.27 m×0.61 m) supplied with treated seawater. To minimize the interference of environmental microbiota on intestinal microbial composition, the treated seawater used in all temperature groups was sourced from the same batch of filtered and disinfected seawater, ensuring consistent initial microbial composition across groups. Each tank maintained a water depth of approximately 30 cm, with PVC tubes placed at the bottom to allow crabs to burrow. The crabs were fed ice fish daily at 17:00, with the feeding amount set at 3–10% of their body weight. After each feeding, uneaten feed in each tank was removed daily two hours later. During the experiment, the water was changed every 5 days, with 1/2 volume of water changed each time. The water quality parameters (salinity 26±2 ppt, ammonia-N<0.5 mg L-1; nitrite<0.10 mg L-1; DO>5 mg L-1 and pH 7.0-9.0) were monitored once a week throughout the experimental period. Measurement methods were as follows: salinity was determined using an optical salinometer (YDJ-lzq, Chuangjimei, China ); Ammonia-N and nitrite concentrations were measured via the Nessler’s reagent spectrophotometric method and N-(1-naphthyl)-ethylenediamine spectrophotometric method, respectively; Dissolved oxygen (DO) was detected with a handheld dissolved oxygen meter (AR8406, Smart Sensor, China); pH was measured using a digital pH meter (PHB-5, Leica, China).
Experimental design
Sample collection
A total of 120 healthy female crabs were randomly and equally assigned to four groups. Four temperature treatments (20℃, 23℃, 26℃, and 29℃) were established, with each treatment consisting of thirty individually housed crabs in separate tanks. The temperature during the experiment was controlled by the use of a heating device. The experiment concluded after 50 days of temperature treatments. At the end of the experiment, nine crabs were randomly sampled from each treatment, weighed, and then these crabs were anesthetized on ice and dissected to obtain intestines. The intestines of each crab were divided into two sections: one was fixed in 4% paraformaldehyde (PFA) for histological analysis, and the other was snap-frozen in liquid nitrogen for microflora analysis.
Histological analysis
The preparation of intestinal sections followed the method described by Lu et al.20 Intestinal tissues were fixed in 4% paraformaldehyde (PFA) at 4℃ for 24 hours, then dehydrated through a series of ethanol solutions with increasing concentrations (70%, 80%, 90%, and 100%). Subsequent clearing was performed. Tissues were then infiltrated with paraffin at 60℃ and embedded in paraffin blocks. The embedded blocks were sectioned into 6μm-thick slices using a rotary microtome (YD-1508R, Jinhua Yidi Medical Equipment Co., Ltd.). The sections were then dewaxed in xylene, followed by rehydration in a series of ethanol solutions with decreasing concentrations (100%, 90%, 80%, 70%) for 5 minutes each, and subsequently rinsed with distilled water. Hematoxylin staining was performed for 5 minutes, followed by differentiation in 1% hydrochloric acid-ethanol for 30 seconds and bluing under tap water for 5 minutes. Then, eosin staining was conducted for 2 minutes. After that, the sections were dehydrated again through ethanol and cleared in xylene. Finally, the sections were mounted with neutral balsam and air-dried. Sections were observed under a light microscope (Nikon SMZ1500, Leica Microsystems) with a 10× objective for overview and 40× objective for measuring intestinal duplicature parameters. Intestinal sections from three crabs were examined in each temperature group. Following Lu et al.20’s method, three intestinal images per group from light microscopy were analyzed using NIS-Elements software to measure intestinal duplicature length and width.
Intestinal microbiota analysis
The DNA of intestinal microorganisms was extracted using the MagPure Soil DNA LQ Kit (Magan) according to the manufacturer’s protocols. DNA concentration was verified by NanoDrop and agarose gel electrophoresis. The genomic DNA was used as a template for PCR amplification with the barcoded primers and Tks Gflex DNA Polymerase (Takara). The V3-V4 regions of the bacterial 16S rDNA were amplified with barcoded primers 343F (TACGGRAGGCAGCAG) and 798R (AGGGTATCTAATCCT). High-throughput sequencing of 16S rRNA was performed using MiSeq (Illumina, San Diego, CA, USA) at Shanghai OE Biotech Co., Ltd. (Shanghai, China).
Raw sequencing data were preprocessed using Cutadapt software to identify and trim adapter sequences. Following trimming, paired-end reads underwent quality filtering, denoising, merging, and chimera removal using DADA2 with default QIIME2 parameters.21,22 Finally, the software generated representative reads and an ASV abundance table. Representative reads for each ASV were selected using the QIIME 2 package. The taxonomic annotation methods are as follows: Bacterial taxa (from phylum to species level) were annotated by BLAST alignment against the Silva database (Version 138); Fungal taxa were primarily annotated at the genus and species levels via the UNITE database (16S/18S/ITS rDNA). All annotations were performed using the q2-feature-classifier plugin with default parameters. Alpha diversity was used to estimate the microbial diversity in intestinal samples, which included the Chao1 index, the Simpson index, the Shannon index, and the Observed Species index.23–26 Alpha indices were analyzed using Mothur 1.30 software. The linear discriminant analysis effect size (LEfSe) method was employed to compare taxonomic abundance profiles. Functional prediction analysis was performed using PICRUSt.27
Statistical analysis
Data statistics and one-way ANOVA were conducted using SPSS 23.0 software (SPSS Inc., USA). Statistical significance was set at P < 0.05. Prior to performing ANOVA, the normality of data was tested using the Shapiro-Wilk test, and the homogeneity of variance was verified via Levene’s test. All data met the assumptions for ANOVA. For post-hoc comparisons, Tukey’s HSD test was applied to determine significant differences between groups. Data are presented as mean ± standard error of the mean (SEM). Then Prism software (GraphPad Software Inc., USA) was used for plotting.
Result
Histology of intestine
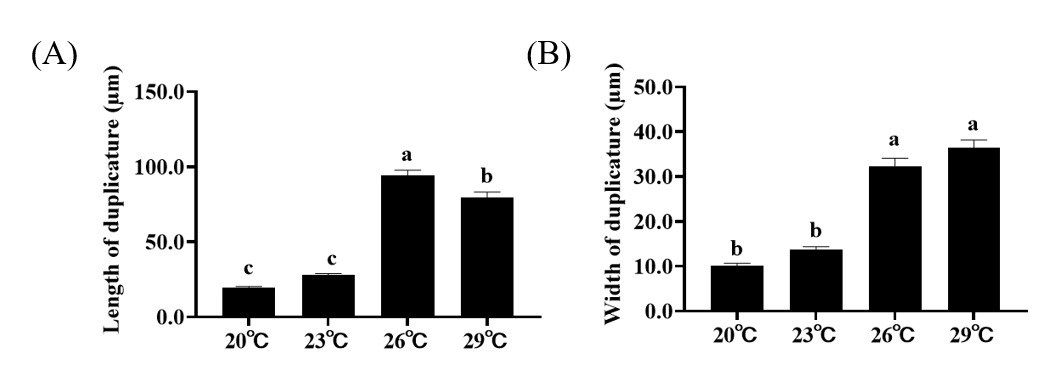
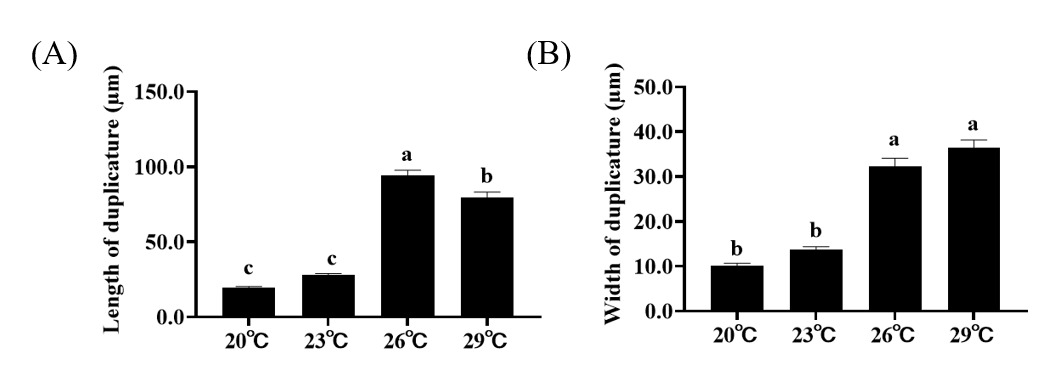
Histological results show that the size of intestinal duplicature gradually becomes larger with the increase of temperature. Further statistical analysis of the measured intestinal histological parameters revealed that the length and width of intestinal duplicature of crabs in higher temperature treatments (26 ℃, 29 ℃) were significantly higher than those of crabs in lower temperature treatments (20℃, 23℃) (Figure 1A, B).
Abundance and diversity of the intestinal microorganisms community
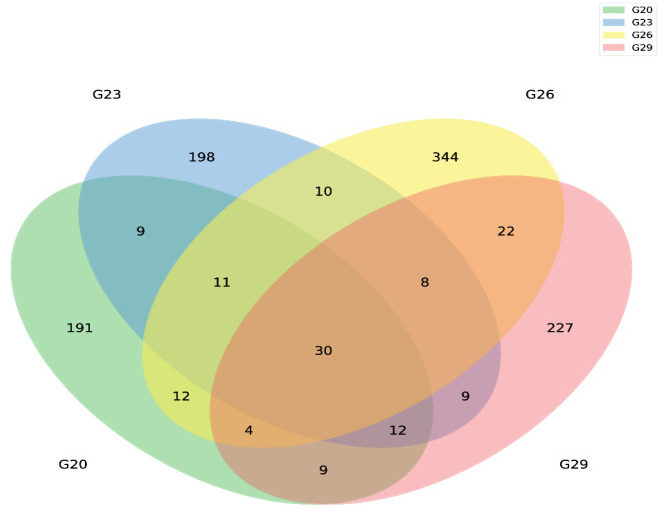
Following sequencing, a total of 956,941 raw reads were obtained from intestinal samples, with an average of 79,745 sequence reads per sample. After chimera removal, 1,096 ASVs were clustered from 863,342 effective tags. A Venn diagram revealed 30 common ASVs across the four groups, along with 191, 198, 344, and 227 unique ASVs at 20℃, 23℃, 26℃, and 29℃, respectively (Figure 2). Alpha diversity analysis of the samples demonstrated that the rarefaction curve approached a plateau, with additional data yielding only a few new ASVs. For alpha diversity indices, the Chao1 and ACE metrics revealed significantly higher species richness at 29℃ compared to the other three temperature treatment groups (Table 1). Moreover, the Shannon and Simpson indices indicated that species diversity at 26℃ and 29℃ was significantly greater than that at 20℃ (Table 1).
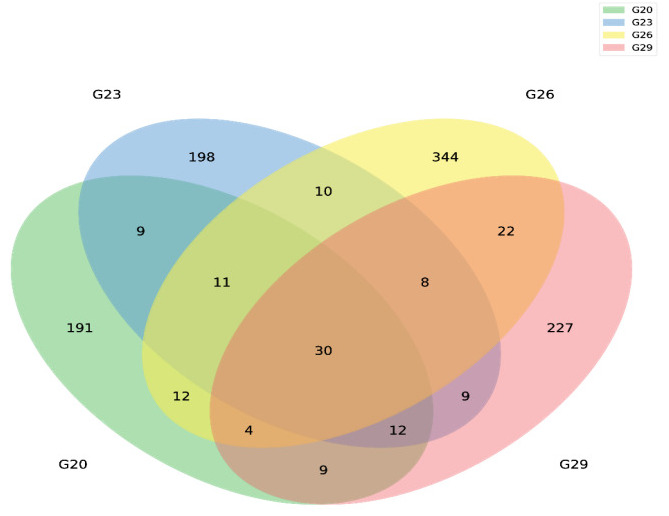
Table 1 Alpha diversity analysis of the intestinal microbiota of P. trituberculatus from different temperatures. Different letters represent the significant difference (p < 0.05) between the groups.
Microbial community composition
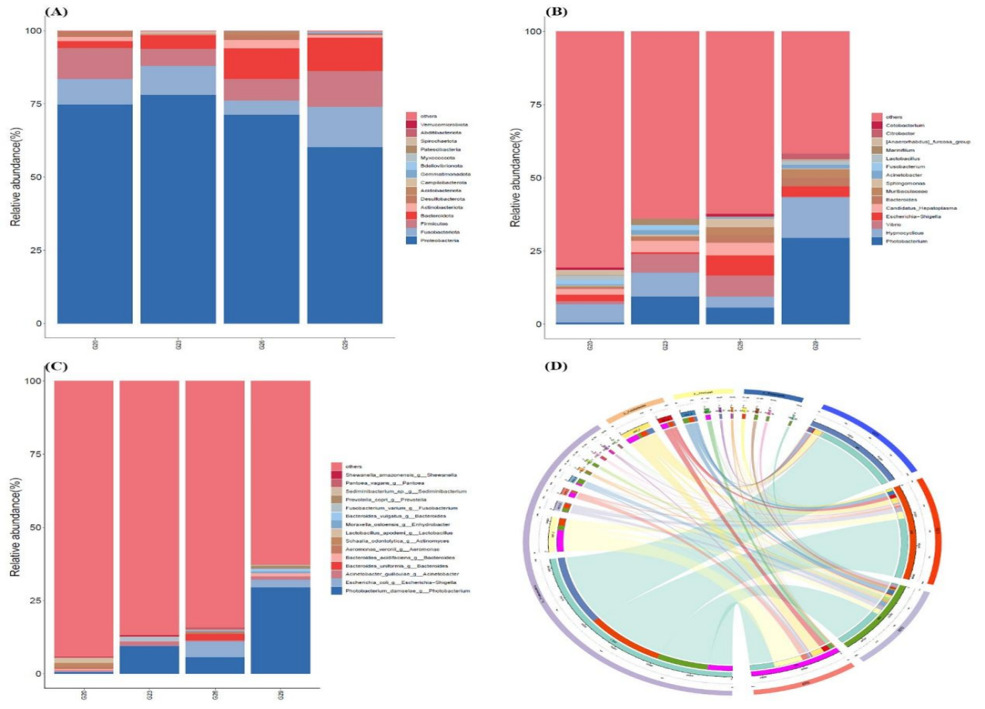
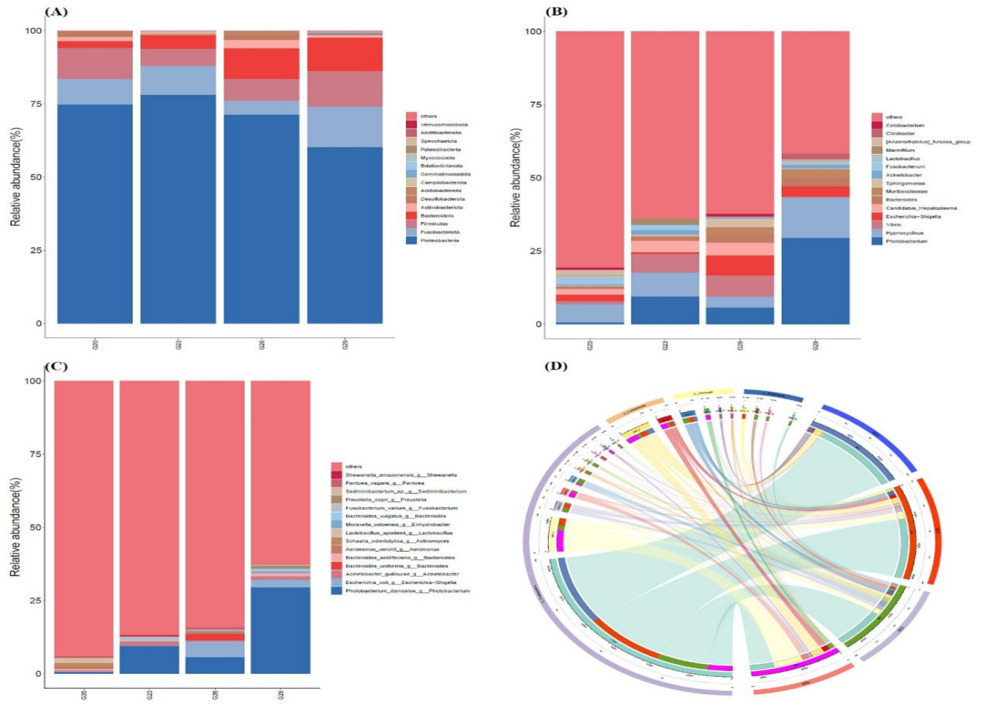
The relative abundance of dominant bacteria varied across different temperature groups. At the phylum level, Proteobacteria, Fusobacteriota, Firmicutes, and Bacteroidota were the major constituents of the intestinal microbial community in P. trituberculatus, with Proteobacteria being the most abundant phylum in all groups (Figure 3A). In the 20℃ group, the intestinal microbial community of P. trituberculatus was dominated by Proteobacteria, Firmicutes, and Fusobacteriota. Compared to the 20℃ group, the proportion of Fusobacteriota decreased significantly in the 26℃ group and increased significantly in the 29℃ group. The proportion of Firmicutes decreased in the 23°C and 26°C groups, while it increased in the 29℃ group. The proportion of Bacteroidota significantly increased with increasing temperature. The proportion of Actinobacteriota was highest in the 26℃ group and lowest in the 23℃ group. At the genus level, the four groups showed distinct bacterial community structures (Figure 3B). The relative abundance of Photobacterium was highest in the 29℃ group (29.57%), but very low in the 20℃ group (0.61%). The relative abundance of Hypnocyclicus was highest in the 29℃ group (13.75%), but very low in the 26℃ group (3.69%). Vibrio showed high abundance in 23℃ and 26℃ groups, occupying approximately 6.35% and 7.14%, respectively, but very low in the 20℃ and 29℃ groups, accounting for just 0.94% and 0.23%. At the species level, Photobacterium_damselae was dominant in the intestinal microbial community of crabs in the 29℃ group (Figure 3C). The relative abundance of Candidatus_Hepatoplasma accounted for a higher proportion in 20℃, 23℃ and 26℃ groups, which was essentially non-existent in the 29℃ group (Figure 3C). Moreover, the proportion of Escherichia-Shigella in the 26 and 29℃ groups was higher than that of the 20 and 23℃ groups.
Changes in the microbial community phylotypes
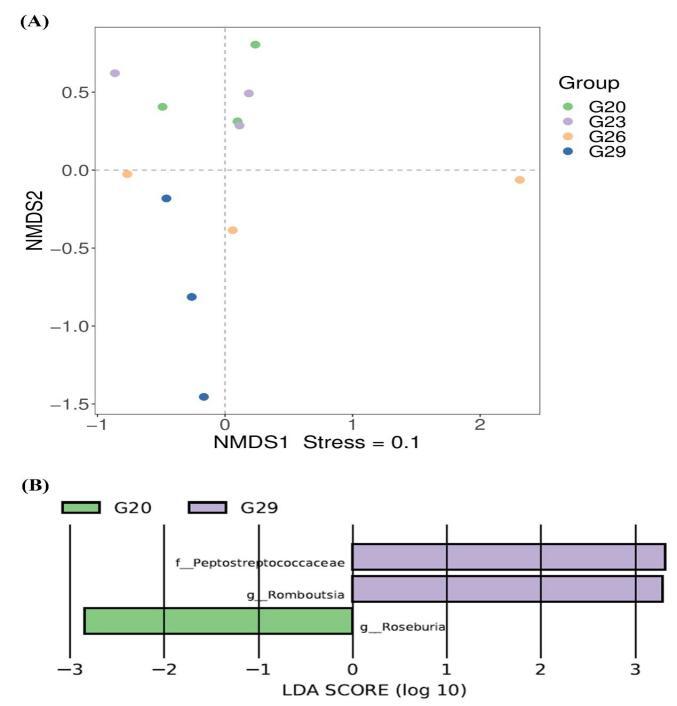
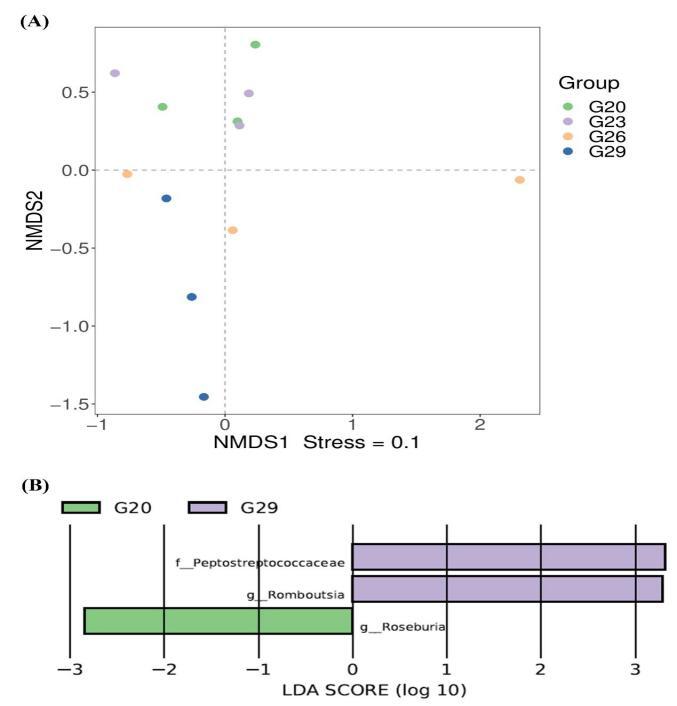
Significant differences in intestinal bacterial community composition were found between the higher temperature group (26℃, 29℃) and lower temperature groups (20℃, 23℃) (Figure 3D). NMDS showed that there was an intersection between 20℃ and 23℃ groups (Figure 4A). Similarly, there was an intersection between 26℃ and 29℃ groups (Figure 4A). In particular, they were distributed in different areas without intersection between the higher temperature group (26℃, 29℃) and the lower temperature groups (20℃, 23℃). To explore the differential abundances of bacterial taxa across temperature treatments, linear discriminant analysis (LDA) was performed to assess the effect size among the 20℃, 23℃, 26℃, and 29℃ groups (Figure 4B). LEfSe analysis revealed greater dissimilarity in the intestinal microbial communities of P. trituberculatus between the 20℃ and 29℃ groups. At the genus level, Roseburia exhibited higher relative abundance at 20℃ than at 29℃, while Romboutsia showed the opposite pattern (higher abundance at 29℃ than at 20℃). At the family level, Peptostreptococcaceae had significantly higher relative abundance at 29℃ compared to 20℃.
Prediction of intestinal microbial function
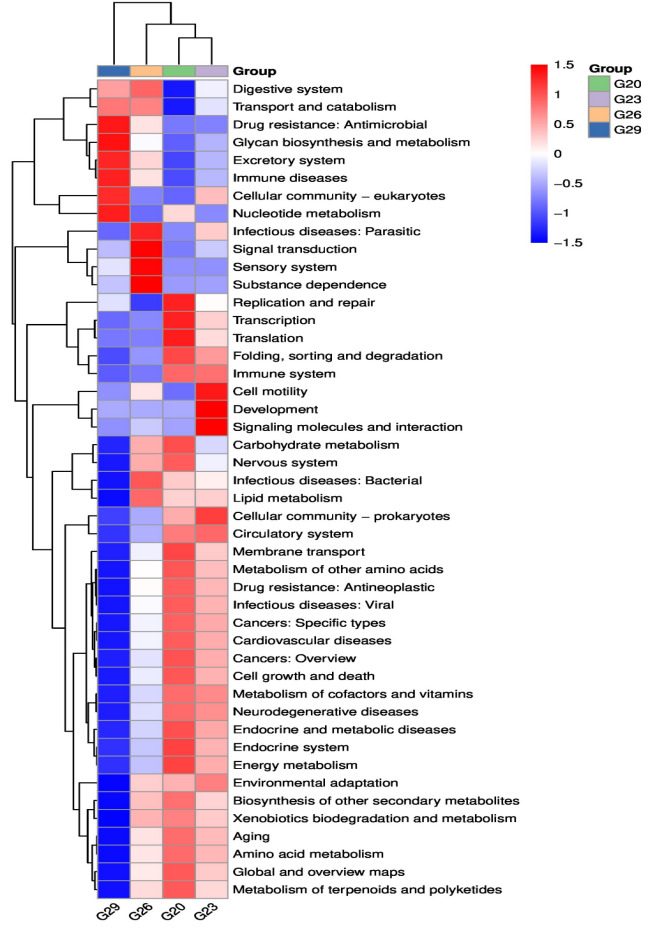
Functional metabolic pathways of the intestinal microbial community were predicted from 16S rRNA gene sequences using PICRUSt2. A total of 46 functional pathways were predicted, and the three most enriched pathways were Carbohydrate metabolism, Amino acid metabolism, and Metabolism of cofactors and vitamins (Figure 5). Specifically, most of the pathways enriched in the 29℃ group were significantly lower than those in the 20℃ group, especially Replication and repair, Carbohydrate metabolism, and Endocrine system. However, some pathways enriched in the 29℃ group were significantly higher than those in the 20℃ group, such as Digestive system, Transport and catabolism, Drug resistance: Antimicrobial, Glycan biosynthesis and metabolism and Immune diseases. Moreover, the KEGG pathways involved in the digestive system, Signal transduction and Infectious diseases: bacterial increased in the 26℃ group compared with the 23℃ group, while the pathways involved in Cell motility, Development and Signaling molecules and interaction exhibited an opposite trend.
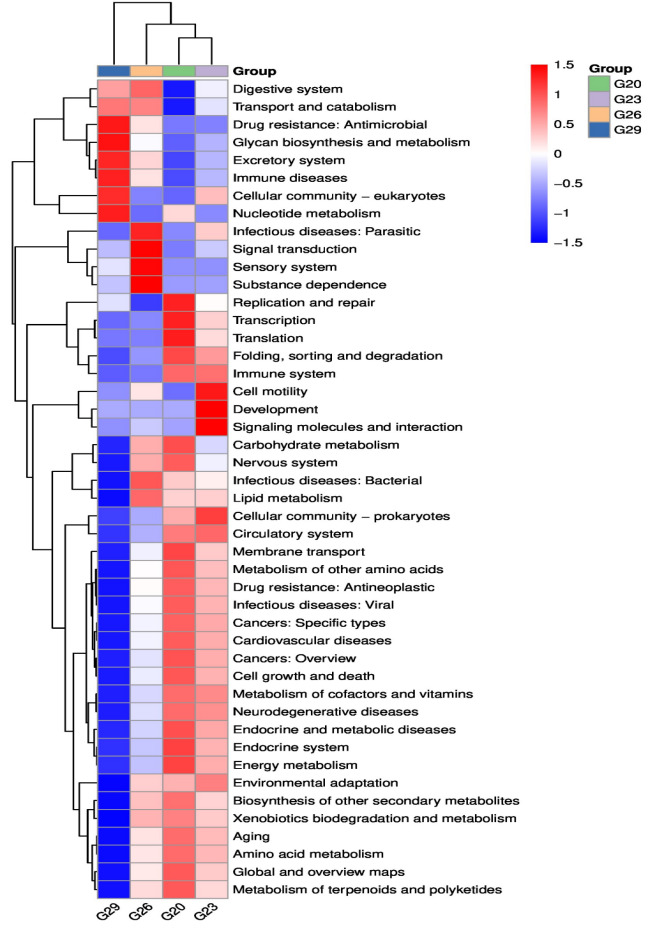
Figure 5 KEGG enrichment analysis of the intestinal microbiota of P. trituberculatus at different temperatures. Red indicates a higher relative abundance of species, and blue indicates a lower relative abundance.
Discussion
In crustacean animals, the intestine is a vital organ for immune defense and nutrient absorption.28,29 Previous studies showed that the presence of intestinal duplicature greatly increases the surface area of the intestinal lumen, providing a basis for digestive and absorptive functions of the intestine.20,30 In the present study, the length and width of intestinal duplicature of crabs in higher temperature treatments (26℃, 29℃) were significantly higher than those of crabs in lower temperature treatments (20℃, 23℃), indicating that elevated temperatures may promote the ability to digest and absorb nutrients by altering the histological structure of the intestine. This phenomenon may be related to the increased metabolic rate in crustaceans due to elevated temperatures. As metabolic demands rise, the body may adapt by enhancing the digestive and absorptive capacity of the intestine. The expansion of intestinal duplicature could be a response to the need for more efficient nutrient uptake to support the increased metabolic activities. This is consistent with the findings of Pan et al.,18 who observed that increased temperature can enhance the nutrient content in the intestinal tissues of P. trituberculatus. However, such positive effects are likely temperature-dependent. Studies on sturgeons have demonstrated that more severe heat stress can lead to intestinal structural damage and functional impairment.31 Therefore, the beneficial structural changes observed at 26–29℃ in the present study may represent the optimal temperature range for P. trituberculatus, beyond which dysbiosis and tissue damage could occur. Notably, other biotic and abiotic factors have been reported to affect intestinal microbial composition in aquatic invertebrates: dietary variations can alter gut microbial communities by providing distinct nutrient substrates, host health status may disrupt microbial homeostasis, and environmental microbiota can colonize the intestine and shape community structure.32–34 However, in this study, these potential confounding factors were tightly controlled. Thus, the observed changes in intestinal microbiota and histological structure were primarily driven by temperature, and the impacts of other variables were negligible in the present experimental design.
It has been proven that temperature has a significant effect on the physiological and ecological processes of organisms (Garcia-Rueda et al., 2023; Morash et al.35). The results of this study showed that there were obvious differences in the intestinal microbial community structure of P. trituberculatus under different temperature conditions. This can probably be attributed to the fact that temperature variations affect the physiological state of P. trituberculatus, thereby altering the intestinal environment, and in turn influencing the survival and reproduction of intestinal microorganisms. Specifically, higher temperatures may accelerate the metabolic rate of P. trituberculatus, leading to increased nutrient secretion and changes in intestinal pH—factors that can selectively favor the proliferation of certain microbial taxa while inhibiting others.36 For example, higher temperatures may lead to an increase in the metabolic rate of P. trituberculatus, as a result of which environmental factors such as nutrient supply and pH value in the intestine are affected, and further, the microbial community structure is impacted.36 The α-diversity index can reflect the richness and diversity of the intestinal microbial community.37,38 Specifically, the Ace and Chao indices reflect the community richness (higher values of the Ace and Chao indices are indicative of greater community richness.), while the Shannon and Simpson indices reflect the community diversity (the larger the Shannon index, the higher the community diversity; the larger the Simpson index, the lower the community diversity).39 Fu et al.40 reported that the intestinal microbial diversity of pufferfish Takifugu fasciatus is higher under high temperatures than under low temperatures. Wang41 found that the intestinal microbial diversity of zebrafish (Danio rerio) at a water temperature of 28℃ is higher than that at 22℃. Similarly, the present study found that the diversity and abundance of intestinal microorganisms in P. trituberculatus increase with the rise in water temperature. This may be attributed to the enhanced metabolic activities of the host, which provide more nutritional substrates for the growth of microbes.
At the phylum level, Fusobacteriota, Firmicutes, and Bacteroidota were the dominant intestinal microbial communities, which suggests that the three phyla play crucial roles in various physiological and metabolic processes of the crab. Fusobacteriota plays an important role in maintaining the balance of the intestinal microbiota, promoting the growth and development of animals, and enhancing immunity.42 Firmicutes can promote the absorption of nutrients and the growth of animals, while Bacteroidota plays an important role in material degradation and nutrient provision.43,44 The dominant phyla exert substantial influence on the intestinal ecosystem, and specific genera within these phyla also make significant contributions. Romboutsia can regulate the structure of the intestinal microbiota and improve lipid metabolism in animals.45 Some species of Peptostreptococcus can metabolize carbohydrates.46 In the present study, significant differences were detected in the relative abundances of these core microbial taxa between the high-temperature groups (26℃, 29℃) and low-temperature groups (20℃, 23℃). The proportions of Bacteroidota, Romboutsia, and Peptostreptococcus in the high temperature group were significantly higher than those in the low temperature group. This phenomenon likely supports the observed enhancement of digestive and metabolic activity in the high-temperature groups, which is consistent with the results of KEGG analysis revealing pathways related to nutrient transport and catabolism. Notably, a similar trend has been reported in European seabass Dicentrarchus labrax juveniles: the abundance of gut microbiota involved in lipid and carbohydrate metabolism increases under high-temperature environments to meet elevated energy demands.47 This confirms that temperature exerts a conserved regulatory role in shaping functional microbial communities. The growth of organisms is not only promoted by beneficial bacteria but may also be inhibited by harmful bacteria.41,48 Photobacterium damselae is the main pathogen that causes diseases in various aquatic animals.49 Some bacteria in the genus Escherichia-Shigella are pathogenic and may cause intestinal infections or other diseases.50,51 Wang et al.51 reported that the abundance of Photobacterium damselae in the intestines of diseased silver pomfret Pampus argenteus was significantly higher than that in healthy individuals. Suzzi et al.52 found that the abundance of Photobacterium damselae was highest in the eastern striped grunter Pelates sexlineatus hindgut at the sampling site with the highest temperature and significantly lower at the sampling site with lower temperature. In the current study, the abundance of Photobacterium damselae and Escherichia-Shigella in the intestine of P. trituberculatus was significantly higher in the 26℃ and 29℃ treatment groups than in the 20℃ and 23℃ treatment groups, and this finding implies that temperature could serve as a critical determinant influencing the abundance of Photobacterium damselae and Escherichia-Shigella in the intestine of P. trituberculatus. The proliferation of these pathogenic bacteria under high temperatures may stem from weakened host immune suppression or enhanced microbial virulence under thermal stress. In contrast, the low-temperature groups maintained a lower pathogenic load, indicating that while high temperatures promote the growth of beneficial metabolic flora, they may also exacerbate disease risks. This reminds us that during the process of crab farming, it is necessary to pay attention to controlling the water temperature to reduce the growth of harmful bacteria and lower the risk of crabs being infected with disease. The temperature range tested in this study (20–29℃) is highly consistent with the natural and aquaculture environments of P. trituberculatus. The seasonal water temperatures in its natural habitats (such as the Yellow Sea and East China Sea) typically fluctuate from 10°C in winter to 28℃ in summer and may temporarily rise to 29℃ during summer heatwaves.53 The results of this study indicate that temperatures at the upper end of this range (26–29℃), while significantly enhancing the digestive and metabolic efficiency of P. trituberculatus, also lead to a significant increase in the risk of pathogenic bacteria.
In this study, the functional metabolic pathways of the 16S rRNA gene sequences of the intestinal microbial community of P. trituberculatus were predicted, and a total of 46 functional pathways were obtained. Among them, carbohydrate metabolism, amino acid metabolism, and cofactor and vitamin metabolism were the top three most enriched pathways. These functions are consistent with the roles played by microorganisms, such as participating in carbohydrate metabolism and amino acid and protein metabolism.50 Pathway enrichment analyses of the intestinal microbes of P. trituberculatus at different temperatures in this study showed that compared with the low-temperature groups (20℃, 23℃), the high-temperature groups (26℃, 29℃) revealed significant enrichment in pathways related to digestion, nutrient transport, and catabolism, which indicated that under a high-temperature environment, intestinal microorganisms were able to participate more efficiently in food digestion as well as nutrient transport and catabolic metabolism processes, which helped to improve the efficiency of food digestion, and were able to provide more sufficient energy and nutrients for the growth of P. trituberculatus, thus promoting its growth and development. This trend aligns with observations in the California mussel Mytilus californianus, where elevated temperatures (26℃) led to higher protease activity and upregulation of heat shock genes (HSPA12A), indicating an organismal strategy to maintain critical metabolic functions under increased energy demands.54 These findings provide valuable insights into conserved metabolic adaptations across species. Furthermore, the results of this experiment found that the intestinal microorganisms involved in the ‘immune-disease’ pathway showed higher abundance under high temperature, which may suggest that while enhanced digestive metabolism supports growth, the upregulation of immune-related microbes could potentially counteract pathogenic risks—as evidenced by the proliferation of Photobacterium damselae and Escherichia-Shigella observed in the present study. These findings align with those of Fevolden et al.,55 who reported superior immunological parameters in rainbow trout Oncorhynchus mykiss reared at 20℃ compared to those at 14℃, indicating that moderate warming may simultaneously enhance both metabolic efficiency and immune readiness.
Conclusion
The present study investigated the effect of temperature on the intestinal histology and microbiota of P. trituberculatus. Higher temperatures (26℃ and 29℃) were associated with a significant increase in the size of intestinal duplicature, indicating enhanced digestion and absorption. The diversity and abundance of intestinal microbiota also increased with increasing temperature, with Fusobacteriota, Firmicutes, and Bacteroidota dominating. At 29℃, Photobacterium and Bacteroidota were more abundant, which may play a role in lipid metabolism. KEGG function prediction showed that the activity of “digestive system” and “transport and catabolism” pathways was enhanced in the high temperature group, which could help the digestion of food and nutrient metabolism, and promote the growth and absorption of food and nutrients. It is noteworthy that the abundance of potential pathogenic bacteria such as Photobacterium damselae and Escherichia-Shigella increased significantly under high temperature. In conclusion, temperature exerts profound effects on both the microbial community and histological architecture of the intestinal tract in P. trituberculatus. While elevated temperatures enhance digestive potential through microbial and structural adaptations, they concurrently increase pathogenic risks. These findings underscore the necessity of implementing balanced thermal management strategies in aquaculture to optimize growth performance while effectively mitigating disease incidence.
Acknowledgements
This study was funded by two projects (No.42106088 and No.32172993) from the Natural Science Foundation of China, The Major Agricultural Technology Cooperation Plan of Zhejiang Province (2024ZDXT17), The plan project of Zhejiang Marine Fisheries Research Institute (No. HYS-CZ-202407, HYS-CZ-202504). Infrastructure costs were partially supported by the Project of Jiangsu Fisheries Science and Technology (SZ-LYG202029).
Authors’ Contribution
Writing – original draft: Meihui Huo (Lead). Investigation: Gaoyuan Yuan (Lead). Data curation: Qingyuan Liu (Equal), Litao Wan (Equal). Formal Analysis: Tong Li (Lead). Validation: Si Chen (Lead). Supervision: Zhiguo Dong (Lead). Writing – review & editing: Meimei Liu (Lead). Project administration: Jie He (Equal).
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
All animal experiments in this study were approved by the Institutional Animal Care and Use Committee of the school of marine science and Fisheries of Jiangsu Ocean University (Approval Code: 2022-03; Approval Date: March 1, 2022).
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.