1. Introduction
The obscure puffer (Takifugu obscurus) is a commercially significant species extensively cultivated in regions such as Jiangsu Province, China.1 It is highly valued for its high nutritional profile, palatable flesh, and relatively low tetrodotoxin content compared to other pufferfish species.2 Over the past decades, T. obscurus production in China has grown rapidly. Statistics reveal that farmed pufferfish output reached approximately 11,830 tons in 2022—representing a 958% increase from 2005.3 This growth has been attributed to advancements in dietary formulations, processing technologies, and the development of low-toxicity strains.4 Despite these improvements, intensive aquaculture practices have exacerbated challenges associated with high-density culture systems, and frequent disease outbreaks and immunosuppression remain major constraints to the sustainable development of T. obscurus aquaculture.5 Conventional disease management strategies rely heavily on broad-spectrum antibiotics, which compromise the sustainability of T. obscurus farming and create significant food safety risks and antimicrobial resistance. In 2024, the World Health Organization (WHO) emphasized that the prudent use of antibiotics is crucial for ensuring aquaculture sustainability and safeguarding public health.6 Accordingly, the development of eco-friendly and safe alternatives has become an urgent priority.
Innovative feed additives represent a promising strategy to address these challenges. Significant advancements have already been made within the aquaculture industry. Numerous studies have demonstrated that polysaccharide-based additives improve the growth performance of aquatic species and enhance immune function and antioxidant capacity.7 Polysaccharides are high-molecular-weight biopolymers consisting of monosaccharide units linked via glycosidic bonds.8 They are widely distributed natural compounds derived from plant tissues, characterized by environmental sustainability, negligible residues, and diverse bioactivities, making them a focal point in feed additive research. Botanical polysaccharides have been shown to stimulate growth and enhance nonspecific immunity,9 as well as improve hepatic health10 and intestinal barrier integrity.11 However, their application in aquaculture remains relatively limited, and the mechanisms of action and optimal inclusion levels for specific species are not yet fully understood. Lycium barbarum, a traditional medicinal herb, is widely distributed across Asia and has been traditionally used as a functional nutraceutical.12 This plant contains numerous bioactive compounds, such as polysaccharides, flavonoids, and carotenoids. Among these, Lycium barbarum polysaccharides (LBP) are the principal bioactive components responsible for its therapeutic effects.13 Extensive research has demonstrated that LBP exhibits potent antioxidant,14 anti-inflammatory, and anti-tumor activities.15 In fish species such as Lateolabrax maculatus,11 Cyprinus carpio,16 and Luciobarbus capito,14 dietary LBP supplementation has been shown to improve growth performance and enhance disease resistance. It also ameliorates intestinal inflammation11 and regulates lipid metabolism.17 However, the optimal dietary inclusion levels of LBP vary considerably among species, necessitating species-specific evaluation.
During their evolution, teleost fish have developed an intestinal microbiota that plays a critical role in host physiology. Functional microbial strains are positively associated with fish growth.18 LBP have been shown to modulate the gut microbiota, promoting the proliferation of Bacteroidetes and inhibiting pathogenic bacteria,19,20 thereby promoting the production of short-chain fatty acids (SCFAs), These changes can enhance growth rates and disease resistance.21 Although LBP has been preliminarily applied in aquaculture feeds, its effects on T. obscurus remain uninvestigated. The molecular weight, conformational structure, and monosaccharide composition of LBP vary according to their source and extraction method.22 To address these gaps, the present study first characterized the monosaccharide composition, molecular weight, and conformation of LBP. Subsequently, a 56-day feeding trial was conducted using diets supplemented with 0 , 0.5, 1.0, 1.5, and 2.0 g/kg LBP. The objective of this study was to evaluate the dose-dependent effects of dietary LBP supplementation on growth performance, antioxidant status, immune response, digestive enzyme activities, intestinal histomorphology, and gut microbiota composition in T. obscurus. We hypothesized that dietary LBP supplementation would improve growth performance and physiological health by enhancing antioxidant capacity, intestinal function, and microbial homeostasis.
2. Methods
2.1. Characterization of LBP
In the present study, Lycium barbarum polysaccharides (LBP) were procured as a commercial product (Cat. No. SP9311; Batch No. 2550614003) from Beijing Solarbio Science & Technology Co., Ltd. (Beijing, China). According to the manufacturer’s specifications, the purity of LBP was ≥90% by ultraviolet-visible (UV-Vis) spectrophotometry.
Given that the biological activity of polysaccharides is closely tied to their molecular structure, the LBP used in this study was previously characterized.23 Characterization included analysis of monosaccharide composition and molecular weight distribution, using high-performance anion-exchange chromatography with pulsed amperometric detection (HPAEC-PAD) and high-performance size-exclusion chromatography coupled with multi-angle laser light scattering and refractive index detection (HPSEC-MALLS-RI). The weight-average molecular weight (Mw) of LBP was 50.94 kDa (±16.417%), and the polysaccharide exhibited a compact random-coil conformation. Monosaccharide analysis revealed that LBP predominantly comprised glucose (0.796) and mannose (0.188), with minor proportions of galacturonic acid (0.006), galactose (0.005), glucuronic acid (0.002), and arabinose (0.002). These structural characteristics provide a basis for interpreting the physiological responses observed in T. obscurus.
2.2. Experimental Diets
Five isonitrogenous and isolipidic diets were formulated, including a control diet (LBP-free) and four experimental diets supplemented with Lycium barbarum polysaccharides (LBP) at levels of 0.5, 1.0, 1.5, and 2.0 g/kg (designated as L1, L2, L3, and L4, respectively). Carboxymethyl cellulose was used as a binder to maintain dietary consistency and stability. For diet preparation, all dry ingredients were finely ground, passed through an 80-mesh sieve, and thoroughly mixed. Fish oil was subsequently added, followed by distilled water (approximately 35% of the total diet weight) containing the pre-dissolved LBP, to obtain a homogeneous dough. The mixture was then pelleted into 2.5-mm-diameter pellets using a pellet mill (Pinzheng Equipment Co., Ltd., China). The pellets were dried in a forced-air oven at 65°C for 8 h, vacuum-sealed, and stored at −20°C until use. The ingredient composition and proximate nutrient levels of the experimental diets are presented in Table 1.
2.3. Experimental Fish and Husbandry Management
Juvenile T. obscurus were obtained from Shuangyang Seed Industry Technology Co., Ltd. (Jiangsu, China). The feeding trial was conducted at the Laboratory of Aquatic Animal Nutrition and Feed, Dalian Ocean University (Dalian, China). Prior to trial initiation, the salinity of the rearing water was adjusted to 4‰ using sea salt (Qingdao Hai Zhi Yan Aquarium Technology Co., Ltd.) in accordance with the protocols described by Yang et al.24 The fish were acclimated in 1,500 L tanks and fasted for 3 days, followed by a one-week pre-feeding period with the control diet to facilitate adaptation. During this period, fish were fed to apparent satiety three times daily (9:00, 13:00, and 18:00). Residual feed and feces were siphoned out 30 min post-feeding, and one-third of the water volume was replaced daily while maintaining salinity at 4‰ under continuous aeration.
Following the acclimation period, the fish were fasted for 24 h and anesthetized with 100 mg/L tricaine methane sulfonate (MS-222, buffered to neutrality with NaHCO3).25 A total of 525 healthy, uniform individuals (initial weight: 3.15 ± 0.07 g) were selected and randomly assigned to experimental tanks after measurement of body weight and length. The trial employed a recirculating aquaculture system (RAS) consisting of 150 L tanks, with 35 fish per tank and three replicates per treatment group. During the trial, the recirculation and filtration systems operated continuously; however, approximately one-third of the water volume was replaced daily to maintain water quality, and salinity was monitored and maintained at 4‰ after water replacement. The feeding trial lasted 56 days. Throughout the trial, water quality parameters were monitored daily and maintained at the following levels: actual water temperature 25.0 ± 0.2 °C, pH ~7.7, dissolved oxygen > 6.0 mg/L, nitrite < 0.01 mg/L, and ammonia-nitrogen < 0.20 mg/L. The daily feeding rate was set at 4% of total body weight, with rations adjusted biweekly based on batch weighing.
2.4. Sample Collection
At the conclusion of the 56-day feeding trial, T. obscurus were fasted for 24 h and euthanized with an overdose of NaHCO3-buffered MS-222. All individuals were weighed and measured, with the final weight and body length recorded. At the end of the experiment, all surviving fish from each replicate were counted and weighed to calculate growth performance and survival rate (SR). Based on these whole-tank data, growth indices including weight gain rate (WGR) and specific growth rate (SGR) were determined. Six fish were chosen at random from each replicate after anesthesia in order to collect the liver, intestine, and kidney for hepatosomatic index (HSI) and viscerosomatic index (VSI).
For biochemical analysis, nine fish per tank were randomly selected for blood collection. Blood samples were immediately collected from the caudal vein using a sterile 1-mL syringe (fitted with a 27-gauge needle) pre-coated with heparin sodium. The samples were incubated at 4°C for 30 min, then centrifuged at 3,000 rpm for 10 min. The resulting plasma was immediately frozen in liquid nitrogen and stored at -80°C for subsequent enzymatic analysis. Following blood sampling, liver and intestinal tissues were collected from three randomly selected individuals per tank; these samples were rapidly frozen in liquid nitrogen and stored at -80°C for subsequent tissue enzyme activity assays.
Furthermore, liver tissues from three fish per tank were collected for quantitative PCR (qPCR) analysis. Midgut tissues were also collected from three fish per treatment and fixed in 4% paraformaldehyde for histological examination (assessment of mucosal folds). The fish used for qPCR, histological examination, and tissue enzyme assays were selected from the same individuals sampled for blood and tissue collection. All sampling procedures were performed on sterile ice trays to ensure sample integrity under aseptic conditions.
The proximate composition of the experimental diets was analyzed according to the standard procedures of the Association of Official Analytical Chemists (AOAC). Specifically, crude protein content was determined using the Kjeldahl method. Crude lipid was measured by Soxhlet extraction with petroleum ether. Moisture content was determined by drying the samples at 105°C until a constant weight was achieved.
2.5. Biochemical Assays and Enzymatic Activity
Plasma, hepatic, and intestinal samples were analyzed for biochemical parameters and enzymatic activities using commercial diagnostic kits in strict accordance with the manufacturer’s protocols, with all assays performed based on independent samples. Plasma indices included total cholesterol (TCHO; Cat. No. A111-1-1), triglycerides (TG; Cat. No. A110-1-1), and total protein (TP; Cat. No. A045-2-2), alongside the activities of acid phosphatase (ACP; Cat. No. A060-1-1), alkaline phosphatase (AKP; Cat. No. A059-2-2), and total bile acids (TBA; Cat. No. E003-2-1). Hepatic parameters included total superoxide dismutase (T-SOD; Cat. No. A001-3-2), TP (Cat. No. A045-2-2), malondialdehyde (MDA; Cat. No. A003-1-2), and catalase (CAT; Cat. No. A007-1-1). Intestinal parameters included lipase (LIP; Cat. No. A054-2-1) and amylase (AMS; Cat. No. C016-1-2).
2.6. Intestinal Histological Analysis
Midgut tissues were fixed in 4% paraformaldehyde for 24 h. Following fixation, samples were dehydrated through a graded ethanol series (75%–100%), cleared in xylene, and embedded in paraffin. The paraffin blocks were sectioned at 5 μm using a microtome and stained with hematoxylin and eosin (H&E). Stained sections were imaged using a PANNORAMIC SCAN scanner (3DHISTECH, Hungary) and analyzed with Case Viewer software. For each section, five intact and well-oriented mucosal folds were randomly selected to measure their height and width. The obtained data were then subjected to statistical analysis to assess significant differences between treatments.
2.7. RNA Extraction and Gene Expression Analysis
Total RNA was isolated from the hepatic tissues of T. obscurus using a commercial RNA extraction kit (Tiangen Biotech, Beijing, China). RNA concentration and purity were assessed using an Ultrospec 3100 Pro spectrophotometer (Amersham Bioscience, UK), with the A260/A280 ratio ranging from 1.8 to 2.0. Subsequently, the RNA was reverse-transcribed into cDNA using FastKing gDNA Dispelling RT SuperMix (Tiangen Biotech, Beijing, China). Primer specificity was verified by the presence of a single peak in the melting curve analysis, while amplification efficiency (90%–110%) and linearity (R2 > 0.99) were confirmed using serial dilutions of cDNA.
Quantitative real-time PCR (qPCR) was performed using FastReal qPCR PreMix (SYBR Green; FP217; Tiangen) on a QuantReady K9600 thermal cycler (Hangzhou Suizhen Biotechnology Co., Ltd., Hangzhou, China). The 20-μl reaction mixtures were subjected to 40 cycles of amplification. The thermal profile consisted of initial denaturation at 95°C for 2 min, followed by 40 cycles of denaturation at 95°C for 5 s and annealing/extension at 60°C for 15 s, with fluorescence acquisition at the end of each cycle. A final melting curve analysis was conducted post-amplification to ensure primer specificity. All assays were performed in triplicate, and the relative expression levels of target genes were calculated using the 2-ΔΔCT method, with β-actin used as the internal reference gene. Primer sequences are provided in Table 2.
2.8. Intestinal Microbial Diversity Analysis in Takifugu obscurus
At the end of the feeding trial, Takifugu obscurus were fasted for 24 h, after which three individuals were randomly selected from each treatment group. Under sterile conditions, the fish were dissected, and posterior intestinal samples were collected into sterile centrifuge tubes. Three posterior intestinal samples from the same treatment group were pooled as one composite sample, immediately frozen in liquid nitrogen, and stored at −80 °C until further analysis.
Genomic DNA was extracted from each composite sample using the E.Z.N.A.® Soil DNA Kit (Omega Bio-tek, USA) according to the manufacturer’s protocol. DNA integrity was assessed by 1% (w/v) agarose gel electrophoresis, while DNA concentration and purity were measured using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, USA). The V3–V4 hypervariable region of the bacterial 16S rRNA gene was amplified by PCR using the specific primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). After purification, the PCR products were quantified using a Qubit 3.0 fluorometer (Thermo Fisher Scientific, USA) and normalized to equimolar concentrations for sequencing library construction. The libraries were sequenced on the BGI G99 platform.
Raw sequence data were processed using the Mothur pipeline. High-quality sequences were clustered into operational taxonomic units (OTUs) at a 97% sequence similarity threshold, and chimeric sequences were removed during quality control. Taxonomic classification was performed against the SILVA database with a confidence threshold of 0.8. To normalize sequencing effort among samples, the OTU abundance table was rarefied to 29,963 sequences per sample prior to downstream community composition analysis.
2.9. Statistical Analysis
Experimental data are presented as mean ± standard deviation (SD). Statistical analyses were performed using IBM SPSS Statistics 29 (IBM Corp., Armonk, NY, USA). Data were initially tested for normality and homogeneity of variances. Following verification of these assumptions, significant differences between the experimental groups were determined using the Waller-Duncan multiple range test. Furthermore, to evaluate the relationship between dietary LBP inclusion levels and growth, antioxidant, and Plasma biochemical parameters, a second-order polynomial regression model (y = ax2 + bx + c) was employed to describe dose–response relationships and estimate the optimal dietary LBP requirement for T. obscurus. The threshold for statistical significance was set at P < 0.05.
3. Results
3.1. Growth Performance
Following the 56-day feeding trial, the FBW, WGR, and SGR of T. obscurus in the LBP-supplemented groups initially increased and then decreased as dietary LBP levels increased. Compared with the control group, FBW, WGR, and SGR were significantly elevated in all experimental groups (P < 0.05), with peak values observed in the L2 group (P < 0.05). Conversely, FCR initially decreased and then increased with increasing dietary LBP levels, reaching its lowest value in the L2 group (P < 0.05). Additionally, except for group L1, HSI was significantly lower across all experimental groups compared with the control (P < 0.05). No significant differences in CF or SR were detected among the treatment groups (P > 0.05). Although VSI showed no significant differences among groups, a decreasing trend was observed with increasing LBP levels (P > 0.05) (Table 3).
3.2. Intestinal Histomorphology
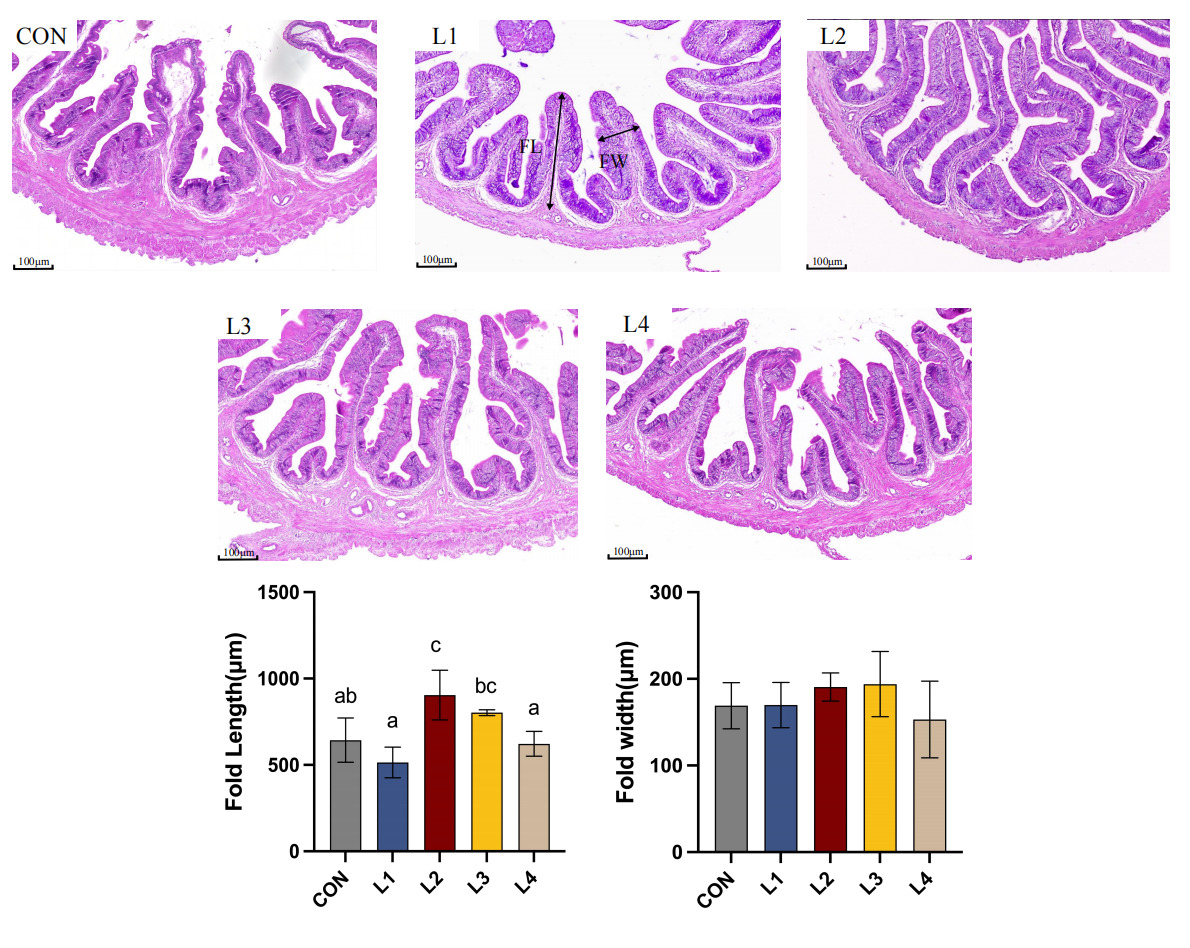
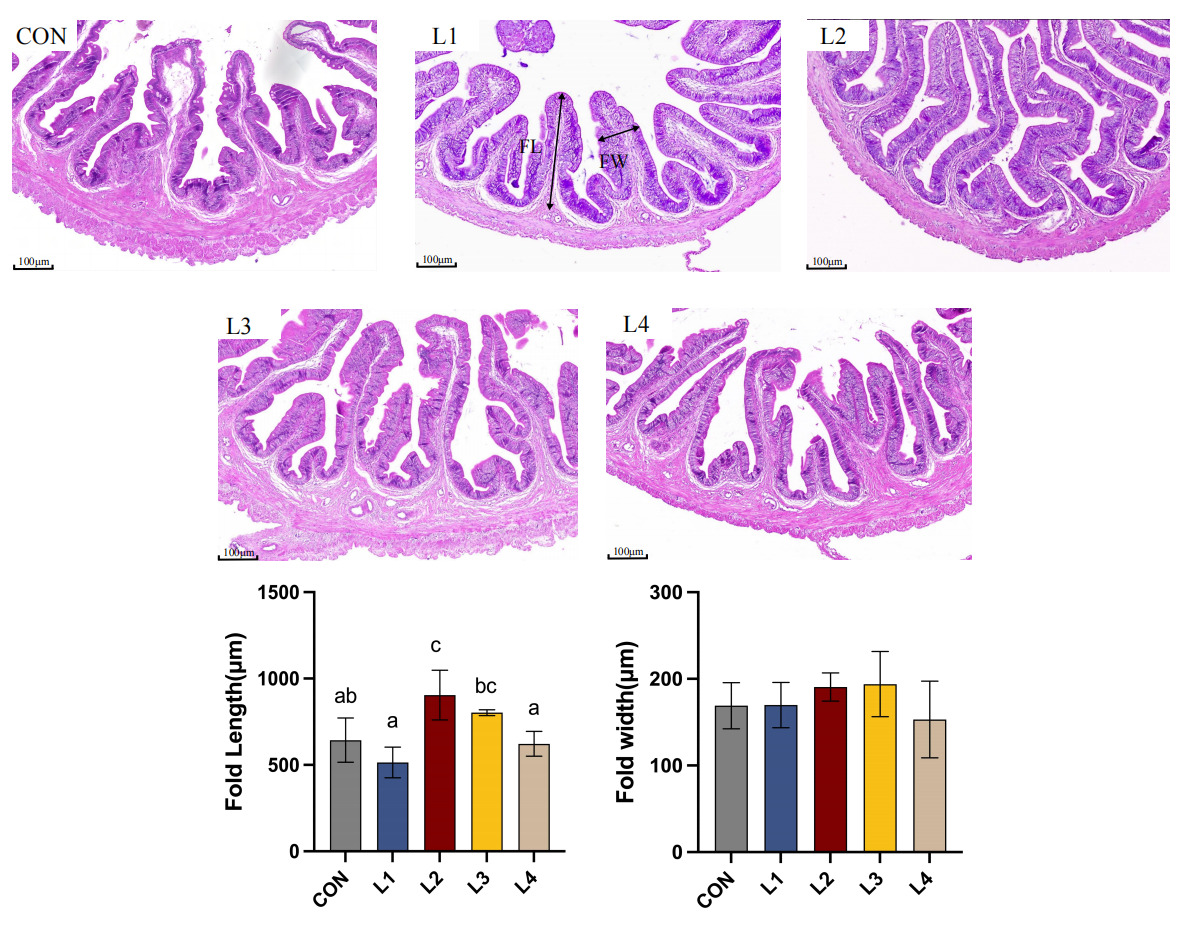
As illustrated in Figure 1, dietary LBP supplementation significantly modulated the intestinal mucosal morphology of T. obscurus. In the control (CON) group, the intestinal mucosal folds appeared relatively short and loosely arranged. In contrast, LBP supplementation enhanced the intestinal architecture, characterized by a more compact and organized tissue structure. The most pronounced improvements were observed in the L2 group, where the fold length (FL) was significantly greater than that in the CON group (p < 0.05). Although no significant difference was observed in fold width (FW) among all groups (p > 0.05), the mucosal folds in the L2 group were more densely distributed and exhibited superior structural integrity.
3.3. Intestinal digestive enzymes
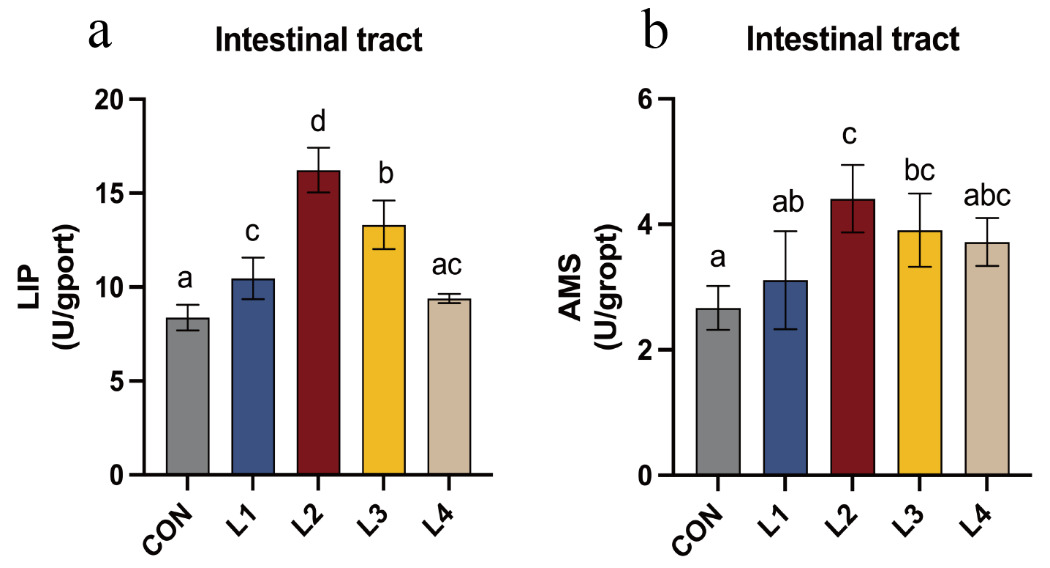
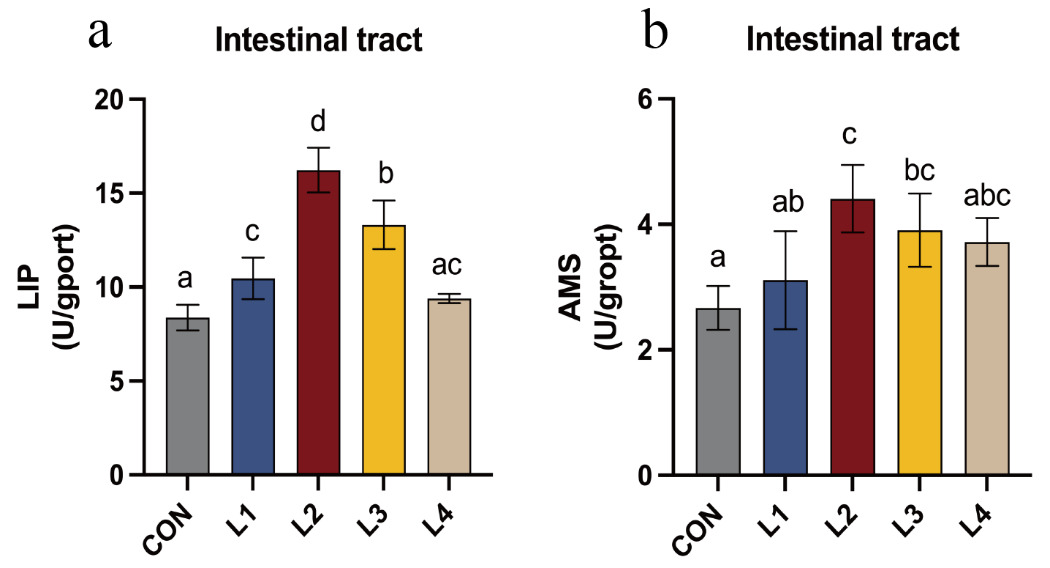
The effects of dietary LBP on intestinal digestive enzyme activities in T. obscurus are shown in Figure 2. LIP activity initially increased and subsequently decreased as dietary LBP levels increased, with the highest value observed in the L2 group. With the exception of the L4 group, LIP activity in all experimental groups was significantly higher than that in the CON group (P < 0.05). Similarly, AMS activity showed a comparable trend, increasing initially and then declining with increasing LBP levels, and reached its maximum in the L2 group. Except for the L1 and L4 groups, AMS activity in the remaining treatment groups was significantly higher than that in the CON group (P < 0.05).
3.4. Plasma biochemical parameters and hepatic antioxidant status
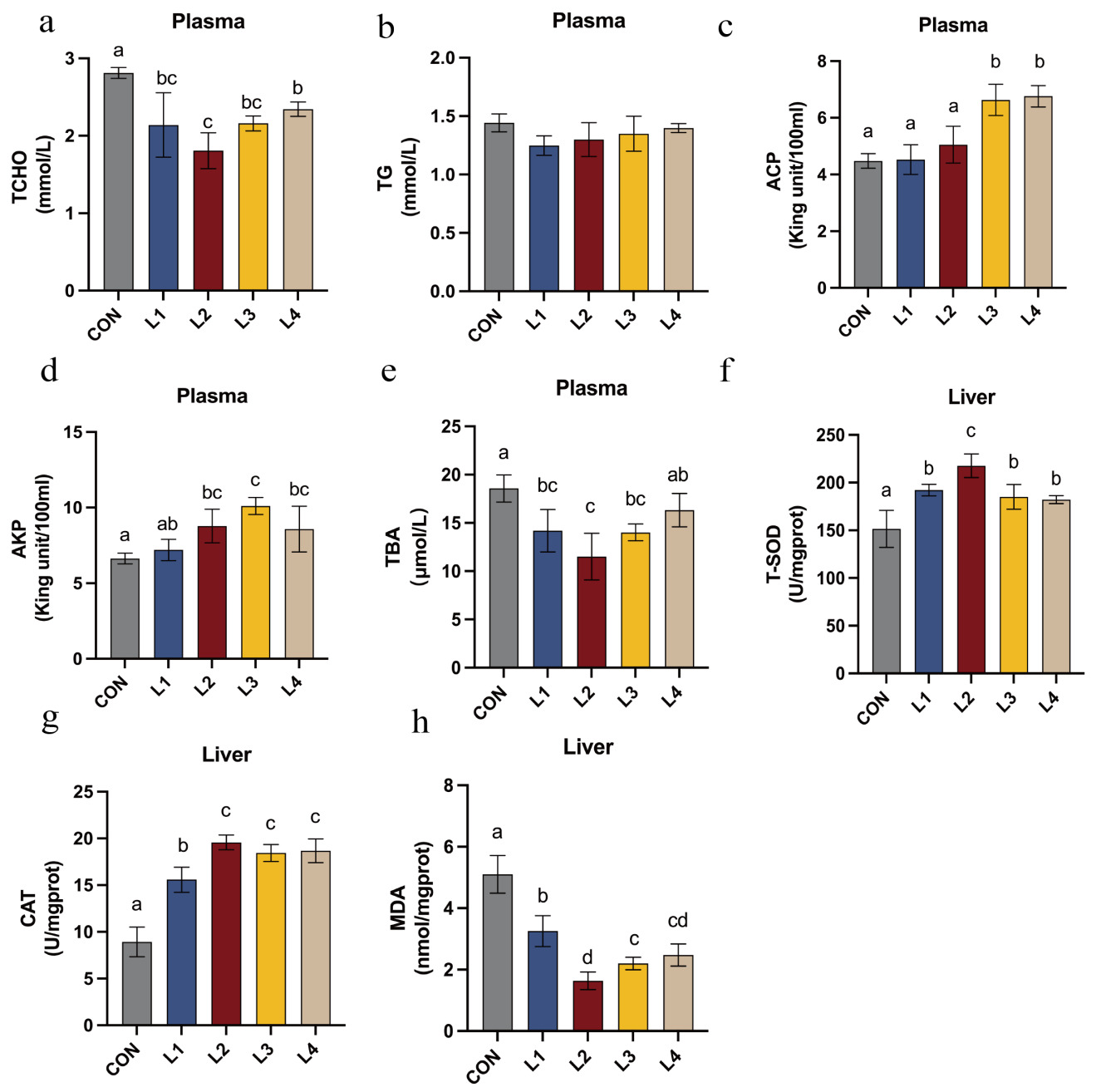
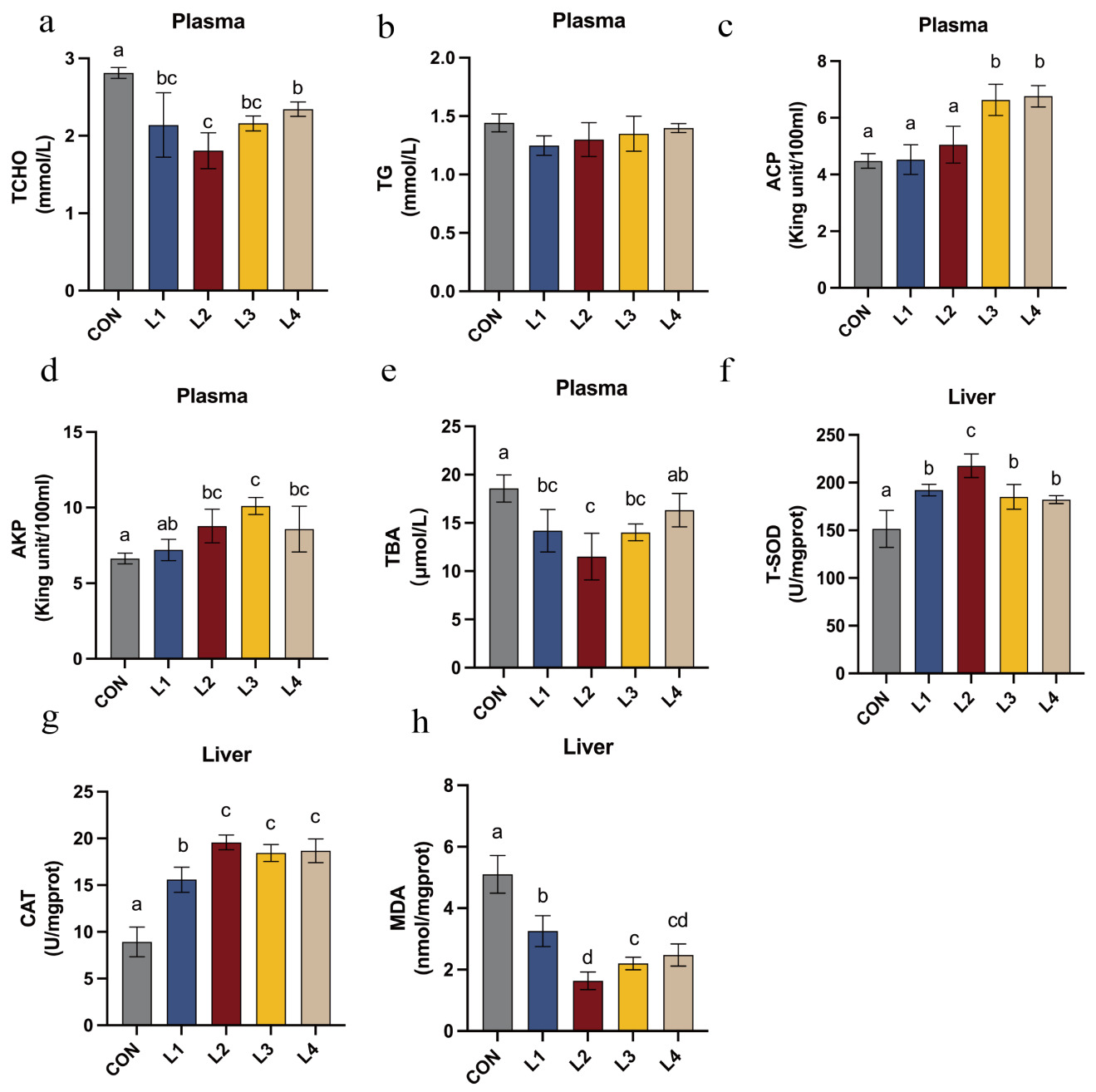
The effects of dietary LBP inclusion levels on the plasma biochemical parameters of T. obscurus are illustrated in Figure 3. Regarding the plasma biochemical profile, levels of TCHO, and TBA were significantly reduced in all LBP-supplemented groups compared with the CON group (P < 0.05). Specifically, TBA and TCHO were lowest in the L2 group and significantly lower than in all other treatment groups (P < 0.05). The activities of ACP and AKP in the L2, L3, and L4 groups were significantly elevated (P < 0.05), with AKP activity reaching its maximum in the L3 group and being significantly higher than those in all other experimental groups (P < 0.05).
In terms of hepatic antioxidant status, LBP-supplemented groups exhibited significantly higher T-SOD and CAT activities, and significantly lower MDA content, compared with the control (P < 0.05). The L2 group exhibited the lowest hepatic MDA levels. Furthermore, T-SOD and CAT activities were highest in the L2 group, significantly exceeding those observed in the remaining treatment groups (P < 0.05).
3.5. Hepatic Gene Expression Analysis
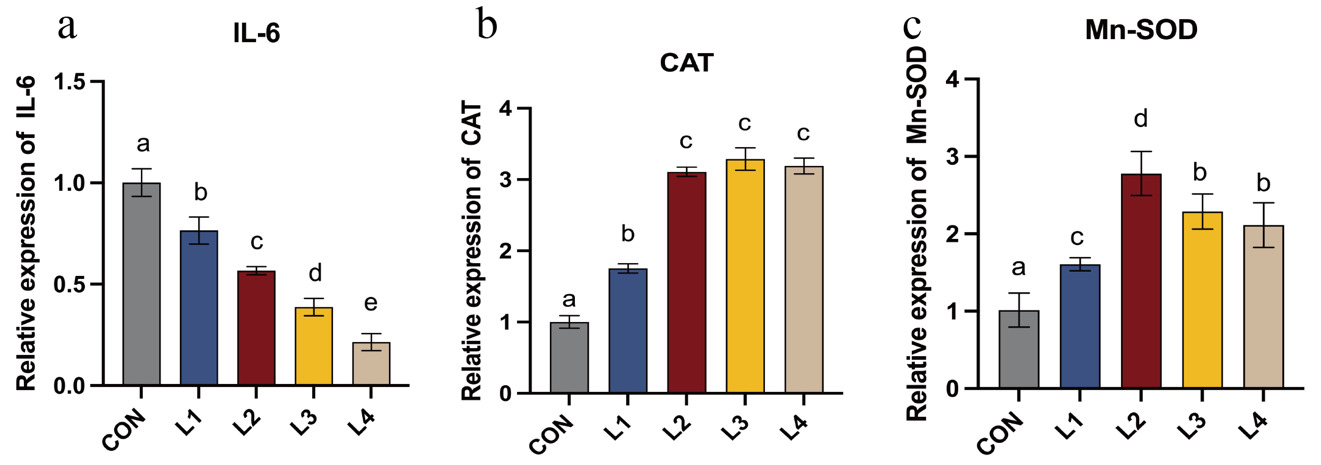
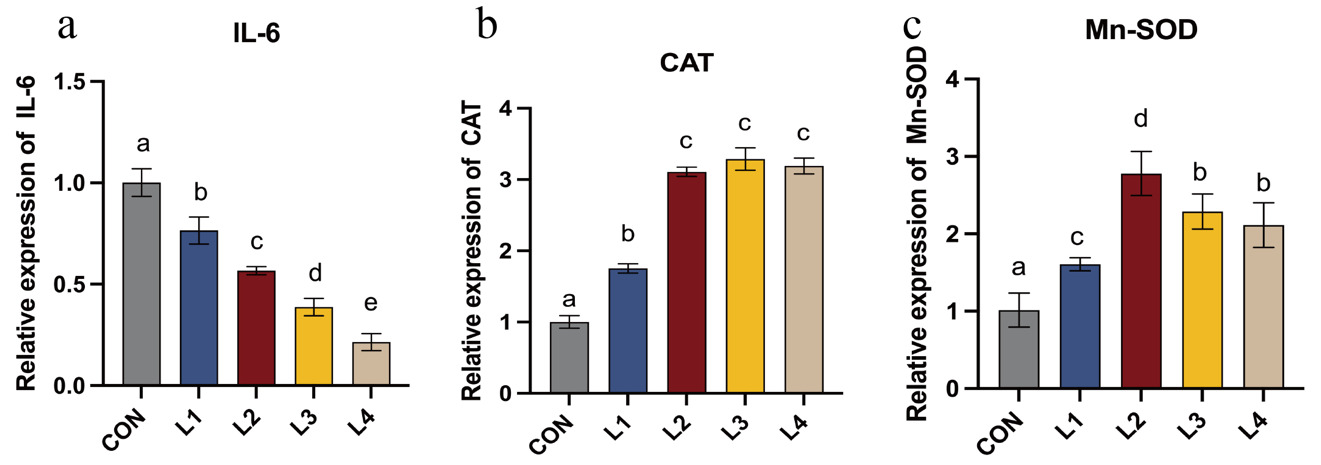
As illustrated in Figure 4, the relative expression levels of antioxidant- and inflammation-related genes in LBP-supplemented groups differed significantly from the control (CON) group. Dietary LBP supplementation induced a progressive downregulation of the pro-inflammatory cytokine IL-6. Significant differences were observed among all treatment groups (P < 0.05), with the lowest expression level observed in the L4 group (P < 0.05). Furthermore, the relative mRNA expression levels of the antioxidant genes Mn-SOD and CAT were significantly higher in all LBP-supplemented groups relative to the control (P < 0.05). Mn-SOD expression increased initially and then decreased with increasing dietary LBP levels, exhibited an initial increase followed by a subsequent decrease, peaking in the L2 group, where it was significantly higher than in all other groups (P < 0.05). While LBP increased CAT expression compared to the CON group, no significant differences were detected among the L2, L3, and L4 groups (P > 0.05).
3.6. Effects of Dietary LBP on Intestinal Microbiota in Takifugu obscurus
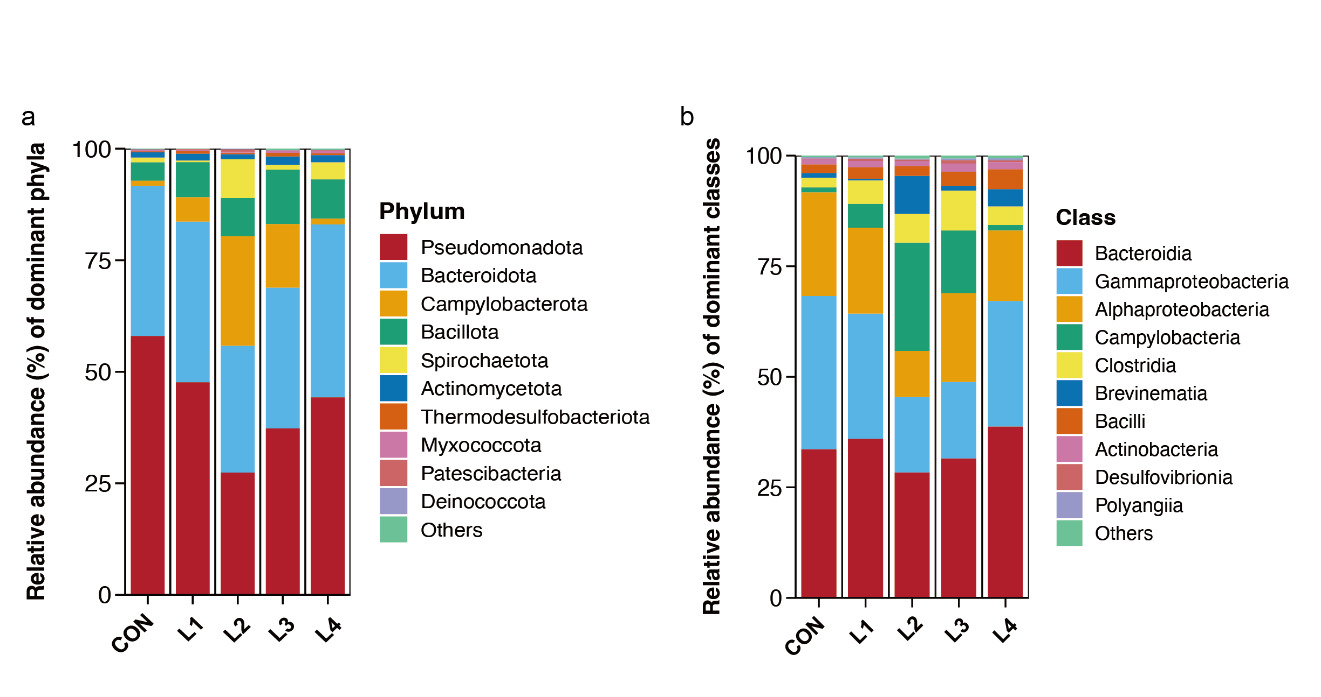
The relative abundances of the top 10 intestinal microbial taxa across different treatment groups (CON, L1, L2, L3, and L4) are presented in Fig. 5.
At the phylum level (Fig. 5a), Bacteroidota and Pseudomonadota were the dominant bacterial phyla across all treatment groups. With increasing dietary LBP supplementation, the relative abundances of Campylobacterota and Spirochaetota initially increased, then declined, reaching their highest levels in the L2 group. Meanwhile, the relative abundance of Pseudomonadota decreased to its lowest level in the L2 group and subsequently increased in the L3 and L4 groups, suggesting that dietary LBP supplementation may modulate the intestinal microbial community structure of Takifugu obscurus.
At the class level (Fig. 5b), Bacteroidia was the dominant bacterial class across all treatment groups. Compared with the CON group, the relative abundances of Gammaproteobacteria and Alphaproteobacteria were markedly reduced in the L2 group, whereas the relative abundance of Campylobacteria increased and reached its highest level.
_and_class_(b)_levels.png)
3.7. Regression Analysis and Estimation of the Optimal Dietary LBP Requirement
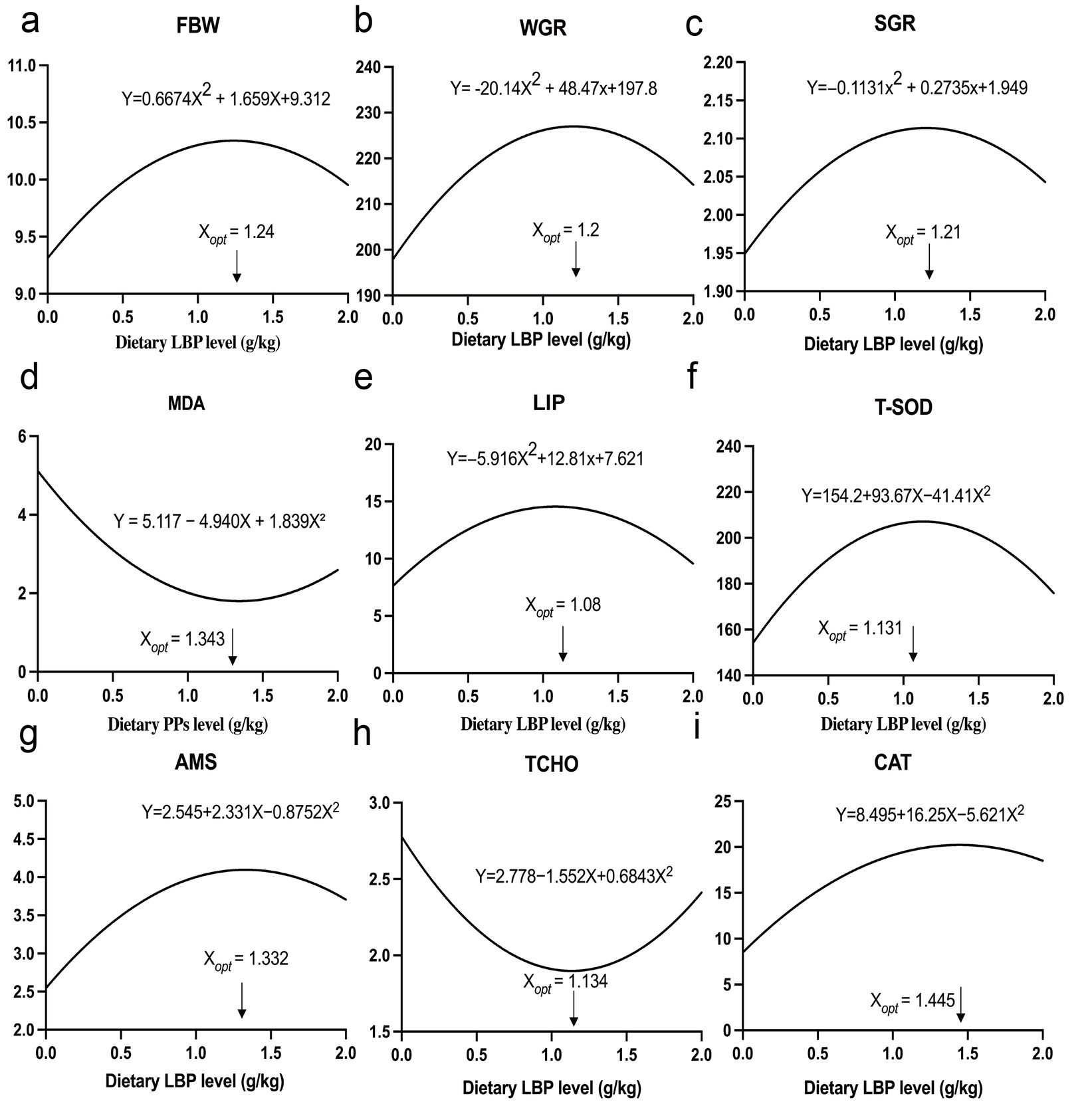
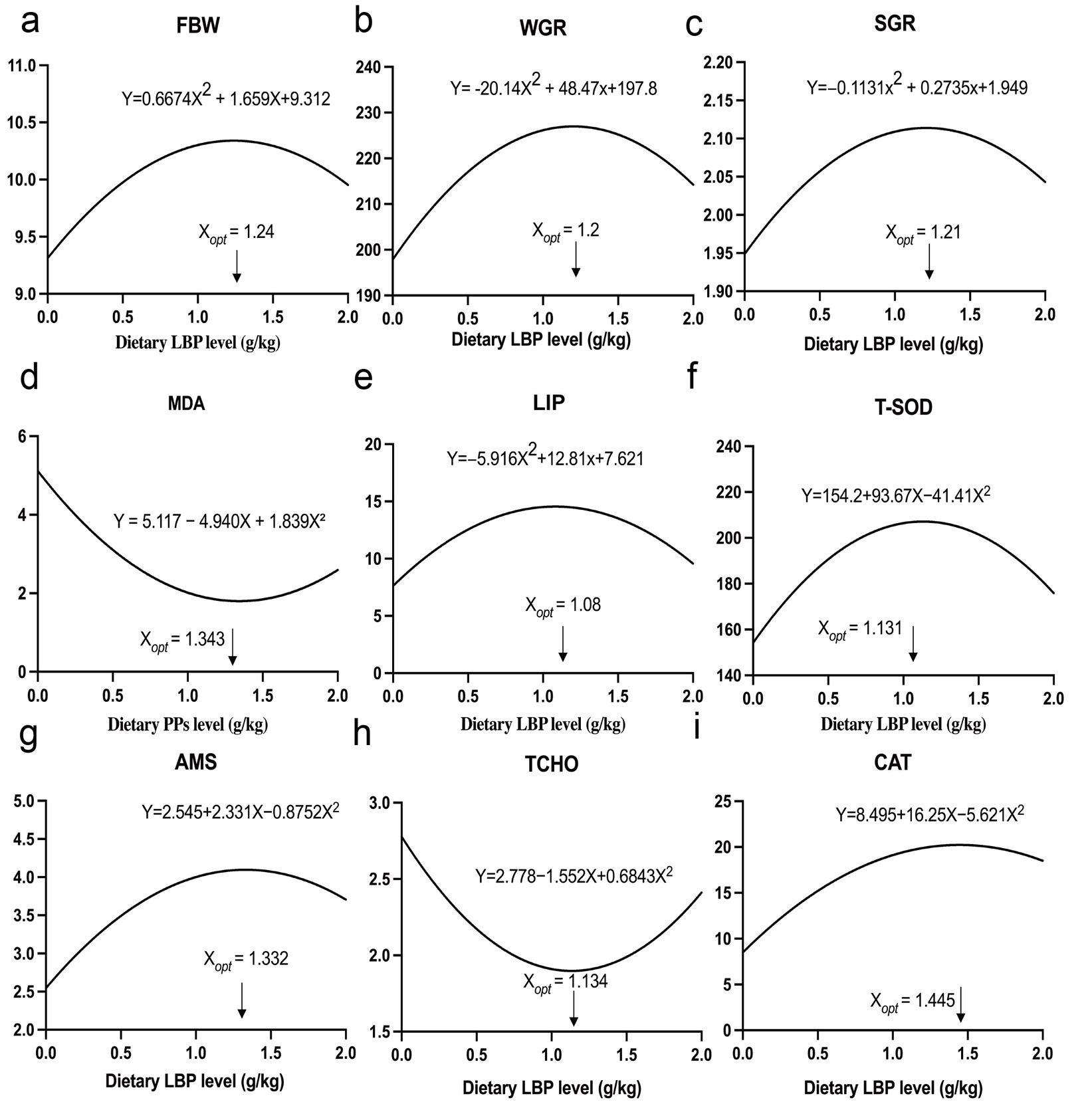
A second-order polynomial regression model (y = ax2+bx+c) was employed to describe the dose–response relationships between dietary LBP levels and the growth performance, antioxidant capacity, and digestive and metabolic parameters of T. obscurus (Fig. 6). The results yielded coefficients of determination (R2) ranging from 0.807 to 0.986, indicating a good fit for all evaluated parameters. Although the L2 group exhibited the most favorable overall physiological performance during empirical observations, regression analysis indicated that the theoretical optimal dietary inclusion levels (Xopt) ranged from 1.08–1.445 g/kg. Considering that no single parameter alone can comprehensively reflect the physiological response of fish to dietary supplementation, the mean value of the nine estimated Xopt values was used as an integrated reference to estimate the dietary LBP requirement. Accordingly, the optimal dietary LBP requirement for T. obscurus was estimated to be 1.235 g/kg.
4. Discussion
Plant-derived polysaccharides have garnered substantial attention across various disciplines due to their potent bioactivity, biodegradability, and lack of harmful residues. As a naturally occurring acidic heteropolysaccharide,29 LBP has been reported to promote growth, enhance antioxidant capacity, and improve immune function.30 Accordingly, LBP are considered promising functional feed additives in animal husbandry and aquaculture. Studies have demonstrated that dietary LBP supplementation in high-soybean-meal diets for Lateolabrax maculatus can partially alleviate hepatic injury and dysregulated glucose-lipid metabolism induced by such diets.31 The biological efficacy of polysaccharides is strongly influenced by their physicochemical properties, particularly molecular weight (MW). In the present study, the measured MW of the LBP was 50.94 kDa. Furthermore, research involving fractionated LBP has indicated that the 46.239 kDa fraction exhibits superior antioxidant activity, free-radical scavenging, and immunomodulatory activities compared with lower-MW fractions.32 Therefore, the LBP used in this study fall within an optimal MW range.
In the present study, dietary LBP supplementation (0.5–2.0 g/kg) significantly improved growth performance and reduced FCR in T. obscurus. These findings are consistent with previous research on other aquatic species, such as Lateolabrax maculatus10 and Hippocampus erectus.33 Similar growth-promoting effects have also been reported for other plant-derived polysaccharides. For instance, laminarin has been shown to improve growth performance and significantly lower the FCR of L. maculatus,34 while mulberry leaf polysaccharides have been reported to promote growth in Micropterus salmoides and mitigate high-starch-induced hepatic injury.7 Growth enhancement is a multifactorial process35 that correlates strongly with antioxidant status, immune function, and intestinal health.36,37 Furthermore, fish growth performance is closely linked to intestinal digestive enzyme activities and villous morphological characteristics.38 Consistent with this, dietary LBP significantly increased lipase (LIP) and amylase (AMS) activities in T. obscurus. Dietary polysaccharides are known to modulate the composition of the gut microbiota, which may stimulate endogenous digestive enzyme secretion. Taxa such as Bacteroidota, Bacillota, and Clostridia possess abundant Polysaccharide Utilization Loci (PULs), which enable them to cleave glycosidic bonds. This degradation process provides a competitive advantage over pathogenic bacteria by enhancing colonization and nutrient utilization.39 Furthermore, Bacillus species, widely used as probiotics in aquaculture, secrete diverse extracellular digestive enzymes that hydrolyze recalcitrant complex carbohydrates.40 Reactive oxygen species (ROS) are inevitable by-products of cellular metabolism; however, excessive accumulation of ROS leads to oxidative stress, which can damage cellular structures and functions.41 The liver is highly susceptible to oxidative stress and is the primary site of synthesis and distribution of key antioxidant enzymes, including CAT and T-SOD.42 Accordingly, the activities of these enzymes reflect the organism’s endogenous antioxidant capacity. Botanical polysaccharides have been shown to augment antioxidant enzyme activity and immune function while directly scavenging free radicals, thereby mitigating oxidative damage and preserving cell membrane integrity.43 This contributes to an overall enhancement of the host antioxidant defense system.44 Consistent with our findings, Luo et al14 reported significant increases in SOD and T-AOC activities following dietary LBP supplementation. However, their study reported a significant reduction in ACP activity, whereas the present study observed no such effect. This discrepancy may be attributed to differences in LBP extraction methods, monosaccharide composition, molecular weight, or species-specific physiological responses. Furthermore, it has been established that polysaccharides can comprehensively enhance immune responses by promoting the development of immune organs, activating macrophages, and inducing dendritic cell maturation.45
Plasma biochemical parameters are key indicators of physiological status, with TCHO reflecting systemic lipid metabolism. In the present study, dietary LBP supplementation significantly reduced plasma TCHO and TBA levels, suggesting that LBP positively modulates lipid metabolism in T. obscurus. These findings are consistent with previous reports.46,47 In contrast, Huang et al17 reported that dietary LBP improved hepatic lipid metabolism in L. maculatus but did not significantly affect Plasma lipid levels. Such discrepancies likely arise from the physicochemical heterogeneity of LBP and interspecies physiological variations. Pseudomonadota was the dominant bacterial phylum in the intestine of T. obscurus , and its relative abundance decreased markedly following dietary LBP supplementation, reaching the lowest level in the L2 group. Previous studies have suggested that an abnormal expansion of Pseudomonadota is a potential microbial signature of intestinal dysbiosis and disease risk,48 whereas a reduction in its abundance may reflect improved microbial stability and intestinal homeostasis,49 In contrast, Bacillota exhibited a slight increase in the LBP-fed groups. Members of Bacillota are known to ferment dietary carbohydrates and produce SCFAs,50 which contribute to intestinal barrier maintenance and host health.51 At the class level, dietary LBP supplementation markedly altered the intestinal microbial community of T. obscurus. Notably, the relative abundance of Gammaproteobacteria decreased substantially in the L2 and L3 groups compared with the control group. Gammaproteobacteria comprise a large proportion of the phylum Pseudomonadota and include numerous opportunistic pathogens, such as species of Aeromonas, Pseudomonas, and Vibrio.52 Previous studies have suggested that the overrepresentation of Gammaproteobacteria is frequently associated with intestinal dysbiosis and compromised intestinal health.53 In contrast, the relative abundance of Clostridia increased in the LBP-fed groups, particularly in the L2 and L3 groups. Members of Clostridia are recognized as important anaerobic fermentative bacteria capable of degrading complex carbohydrates and producing beneficial metabolites, including short-chain fatty acids (SCFAs).54 These metabolites contribute to intestinal barrier integrity, immune regulation, and nutrient utilization.51 Therefore, the microbial shifts observed in the present study may indicate that dietary LBP promoted a more favorable intestinal microbial environment. These changes may partly explain the improvements in intestinal morphology and growth performance observed in fish fed the LBP-supplemented diets.
Mechanistically, the ability of polysaccharides to inhibit lipid absorption depends on their steric conformation and hydrophobicity. Distinct extraction protocols may alter the bile acid sequestration capacity of LBP,55,56 thereby affecting lipid absorption processes in the intestine. While it is conventionally accepted that higher molecular weight and viscosity enhance the lipid-blocking potential of polysaccharides,56 recent evidence suggests that molecular weight is not the sole determinant of bile acid-binding affinity. Low-molecular-weight polysaccharides may possess a larger specific surface area, facilitating increased interaction with bile acids.57 Furthermore, variations in fish species and developmental stages result in distinct capacities for degrading and assimilating macromolecular polysaccharides. Given the complex interplay among molecular weight, conformation, and monosaccharide composition, further studies are needed to elucidate the structure–activity relationships of LBP. It should also be noted that dietary LBP supplementation was achieved by replacing an equivalent amount of carboxymethyl cellulose, which was used as a feed binder in the experimental diets. However, the maximum replacement level was only 0.2%, and therefore, the minor variation in CMC content among treatments was unlikely to have materially influenced the observed physiological responses. In summary, four dietary LBP inclusion levels (0.5, 1.0, 1.5, and 2.0 g/kg) were evaluated in this study. While observational data suggested 1.0 g/kg as the optimal concentration for growth performance, a comprehensive second-order polynomial regression analysis—integrating growth, enzymatic activities, and other physiological indices—indicated that the optimal dietary LBP requirement for T. obscurus is 1.235 g/kg. It should be noted that gut microbiota analysis was conducted on a single composite sample per dietary treatment. Therefore, the microbiota results should be regarded as preliminary and exploratory observations of community composition rather than statistically supported differences in microbial diversity. Future studies incorporating biological replicates and comprehensive diversity analyses will be necessary to further validate the effects of dietary LBP on the intestinal microbiota of T. obscurus.
5. Conclusion
In summary, dietary LBP supplementation significantly enhances the growth performance, intestinal histomorphology, and enzymatic activities of T. obscurus, with the L2 group exhibiting the most pronounced improvements. Regression analysis of the growth and physiological parameters indicates that the theoretical optimal inclusion level of LBP is 1.235 g/kg. These findings suggest that LBP is an effective functional feed additive for improving the growth, immune function, and antioxidant capacity of obscure puffer, providing a robust scientific basis for its integration into sustainable aquaculture practices.
Acknowlegments
This work was supported by the Key Laboratory of Pufferfish Breeding and Culture in Liaoning Province [2021JH13/10200005]; the Dalian Science and Technology Talent Innovation Support Policy Project [2024RG008]; the Dalian Ocean University Science and Technology Innovation Team Project [B202102]; the Liaoning Province Applied Basic Research Program [2025JH2/101300086]; and the Liaoning Provincial Major Science and Technology Project [2025JH1/11700001].
Authors’ Contributions
Conceptualization: Shenglin Yue (Equal), Fuqiang Wang (Equal). Methodology: Shenglin Yue (Equal), Zixiang Lin (Equal). Investigation: Shenglin Yue (Equal), Fuqiang Wang (Equal), Yiyang Huang (Equal), Zixiang Lin (Equal). Resources: Shenglin Yue (Equal), Fuqiang Wang (Equal), Yiyang Huang (Equal), Zixiang Lin (Equal). Formal Analysis: Shenglin Yue (Equal), Zixiang Lin (Equal). Writing – original draft: Shenglin Yue (Equal), Yiyang Huang (Equal), Zixiang Lin (Equal). Validation: Shenglin Yue (Equal), Fuqiang Wang (Equal), Yiyang Huang (Equal), Zixiang Lin (Equal), Yingkang Sun (Equal), Lu Peng, Xiuli Wang (Equal), Yuzhe Han (Equal), Tongjun Ren (Equal), Xiaoran Zhao (Equal). Writing – review & editing: Fuqiang Wang (Equal). Funding acquisition: Fuqiang Wang (Equal), Xiuli Wang (Equal), Kun Li (Equal). Supervision: Fuqiang Wang (Lead). Project administration: Fuqiang Wang (Lead).
Ethical conduct approval – IACUC
This study was approved by the Animal Ethics Committee of Dalian Ocean University (Approval No. 20250026) and conducted in strict accordance with institutional guidelines for the care and use of laboratory animals.
Data Availability
Data can be obtained from the corresponding author
Conflicts of Interest
The authors declare no competing interests.