

1. Introduction
The turbot (Scophthalmus maximus), originally native to Europe, was introduced to China in 1992.1 It is recognized as a crucial commercial fish species due to its rapid growth rate, low-temperature resistance, high feed conversion rate, and extensive industry chain, making it a cornerstone of industrial farming in China.2 Marine aquaculture has experienced rapid development and has transitioned towards intensive factory farming. However, this rapid expansion has also led to frequent disease outbreaks, necessitating increased antibiotic production to prevent disease. Unfortunately, the long-term use of antibiotics has caused numerous adverse effects, including increased antimicrobial resistance, persistent accumulation in organisms, and disruption of microbial equilibrium within the animal gut.3
Given the marine fish farming industry’s focus on green development, it is crucial to seek safe, efficient, environmentally friendly, and residue-free products. Probiotics are widely considered the best solution for ensuring product quality, economic benefits, disease resistance, and immunity in marine fish. Currently, probiotics are globally recognized as natural alternatives to antibiotics due to their high ecological value. The Food and Agriculture Organization (FAO) and the World Health Organization (WHO) define probiotics as “live microorganisms that, when administered in sufficient quantities, provide health benefits to the host.” Practical applications in aquaculture have confirmed the advantages of using probiotics. These advantages include promoting metabolism and growth, improving the micro-ecological structure, increasing feed conversion efficiency, and restoring the aquatic environment. Probiotics achieve these benefits by producing digestive enzymes, secreting antibacterial substances, stimulating immunity, interfering with population sensing, and competing for adhesion sites.4–6
While previous research primarily focuses on probiotics or prebiotics that are not derived from fish, such as Lactobacillus, Bacillus, oligofructose, grobiotic-A, and oregano oil,7–10 these probiotics or prebiotics may not effectively colonize the intestine or dominate the intestinal flora compared to endogenous probiotics.11 In our previous study, we isolated a Halomonas spp. strain from the intestine of healthy turbot and identified it as Halomonas titanicae. H. titanicae was initially isolated and identified from rusticle samples formed by a consortium of microorganisms collected from the RMS Titanic wreck site.12 Halomonas spp. have mainly been studied for ecological applications rather than for aquaculture. However, Halomonas sp. B12, which was isolated from the intestine of shrimp (Fenneropenaeus chinensis), significantly enhanced the shrimp’s resistance to white spot syndrome virus (WSSV).13 Similarly, H. titanicae was also found in wastewater, activated sludge, oil fields, wetlands, and other areas with complex organic compositions, indicating its potential to degrade organic pollution in aquaculture environments.
H. titanicae, being an endogenous microbe, shows promise as a candidate for the development of innovative aquatic probiotics. However, the potential role of H. titanicae has yet to be explored. To determine the suitability of this microbial strain as a probiotic, we conducted a comprehensive examination of its effects on the growth performance, digestibility, antioxidant activity, intestinal histology, and intestinal flora of juvenile turbot. Additionally, we conducted a continuous challenge assay to assess the prolonged probiotic performance of this newly discovered strain after H. titanicae feeding was discontinued.
2. Materials and methods
2.1. Selection of potential probiotic bacteria
2.1.1. Isolation of potential probiotics
In our previous study, we isolated several strains of dominant intestinal bacteria (such as Bacillus velezensis, Pseudoalteromonas issachenkonii, Psychrobacter nivimaris, Bacillus simplex, and Halomonas titanicae) from the healthy intestinal tract of farmed turbot cultured in a fishery in Yantai, Shandong Province, China. The purified strain was stored in MRS medium containing 15% sterile glycerol at −80 °C until use.
2.1.2. Screening of potential probiotics
A pre-experiment was conducted to select a strain with good probiotic properties from these strains. The experimental site and the source of the 30-day-old experimental fish (0.35 ± 0.03g) were the Weihai Aquatic Technology Company in Rongcheng, Shandong Province. These purified strains were stored in MRS medium containing 15% sterile glycerol solution and kept at −80 °C until use. In the pre-experiment, a common feeding group and a group supplemented with the candidate strains were established. These candidate strains were added to the basal diet at 1×1011 CFU/mL. The nutrient and ingredient composition of the basal diet is shown in Table 1. The powdered feed is thoroughly mixed with the probiotic solution and physiological saline. The prepared feeds were naturally dried at room temperature and stored at 4 °C. To ensure the activity of the added strains, fresh diets should be prepared daily. Each treatment consisted of three replicates of 60 fish. The water temperature, pH and salinity were maintained at 17 ℃ ± 1 ℃, 8.0 ± 0.3 and 29.0 ± 0.5, respectively. The fish were fed twice a day (at 9 a.m. and 6 p.m.). On day 30, 20 fish were randomly selected from each group. Their length and weight were measured using vernier caliper and electronic balance.
2.2. Evaluation of probiotic properties of the candidate strains
After a 30-day pre-experiment, the strain with the most pronounced growth-promoting effect on turbot was selected. Subsequently, the strain’s probiotic properties were studied in depth from various perspectives.
2.2.1. Security evaluation and Experimental management
In this experiment, the hemolysis test was employed to evaluate the biological safety of the potential probiotic strain. The strain was streaked onto Columbia blood agar plates containing 5% (v/v) defibrinated sheep blood and incubated at 28 °C under aerobic conditions for 48 h. The absence of clear zones (β-hemolysis) or greenish surrounding areas (α-hemolysis) around colonies indicated γ-hemolysis and a lack of hemolytic activity, confirming the strain’s biosafety for further application.14 The experimental feed configuration, feeding management, and feeding condition were the same as those of the pre-experiment model.
2.2.2. Effect of potential probiotic supplementation on growth index
Twenty fish were randomly selected from each group on day 60. The length and weight were measured using a vernier caliper and an electronic balance.
2.2.3. Effect of potential probiotics supplemented on digestive and immune enzyme activity
On days 30 and 60, nine turbot fish from each group were sampled and dissected on ice using sterile instruments. The liver lobe was selected as the tissue to determine the activities of superoxide dismutase (SOD), catalase (CAT), and alkaline phosphatase (AKP) as indicators of antioxidant activity. Trypsin (TPS), lipase (LPS), and amylase (AMS) were selected to indicate digestive enzyme activity. Tissue samples were homogenized in ice-cold physiological saline (1:9, w/v) and centrifuged at 3,000 × g for 10 min at 4°C. Enzyme activity was determined using a kit from the Nanjing Jiancheng Bioengineering Institute, according to the kit instructions. The mid-gut was also dissected at the same time, frozen in liquid nitrogen, and stored at -80 ℃ for analysis of intestinal flora.
2.2.4. Effect of potential probiotic supplemented on intestinal structure and microbial composition
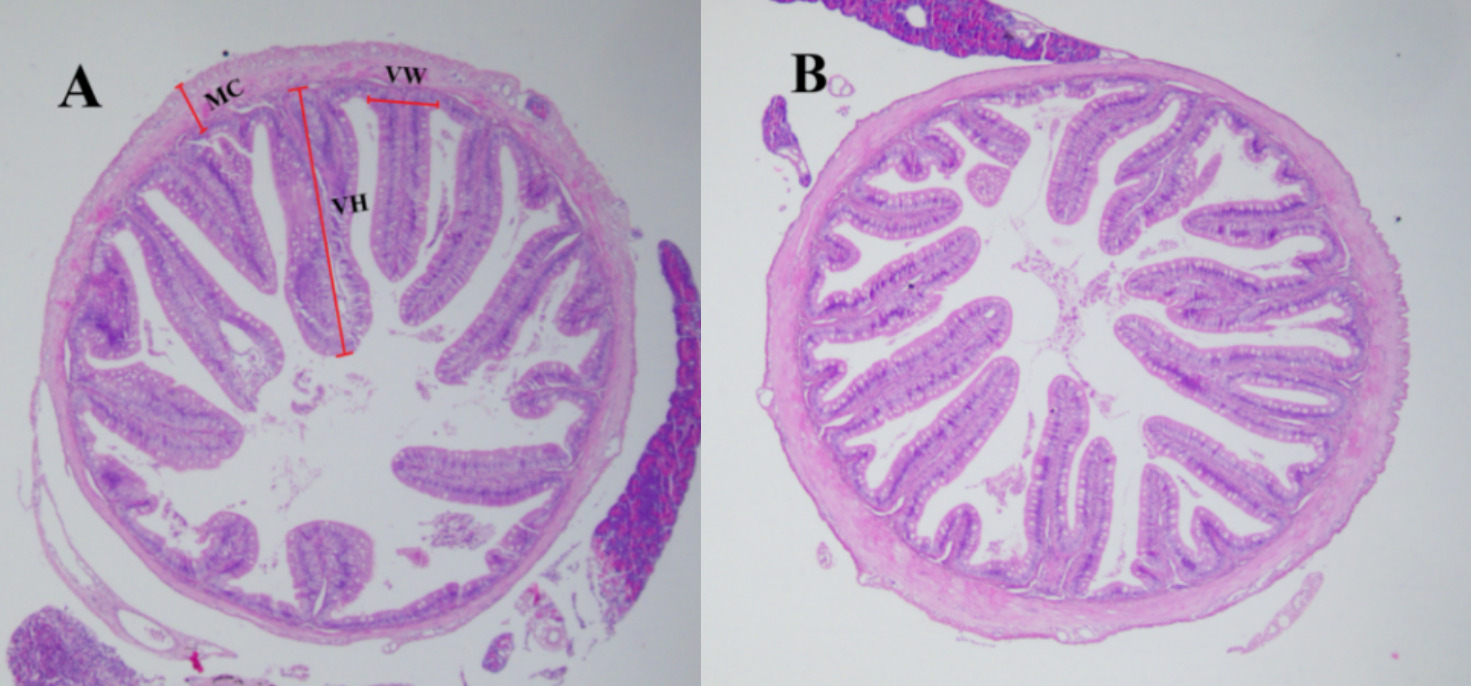
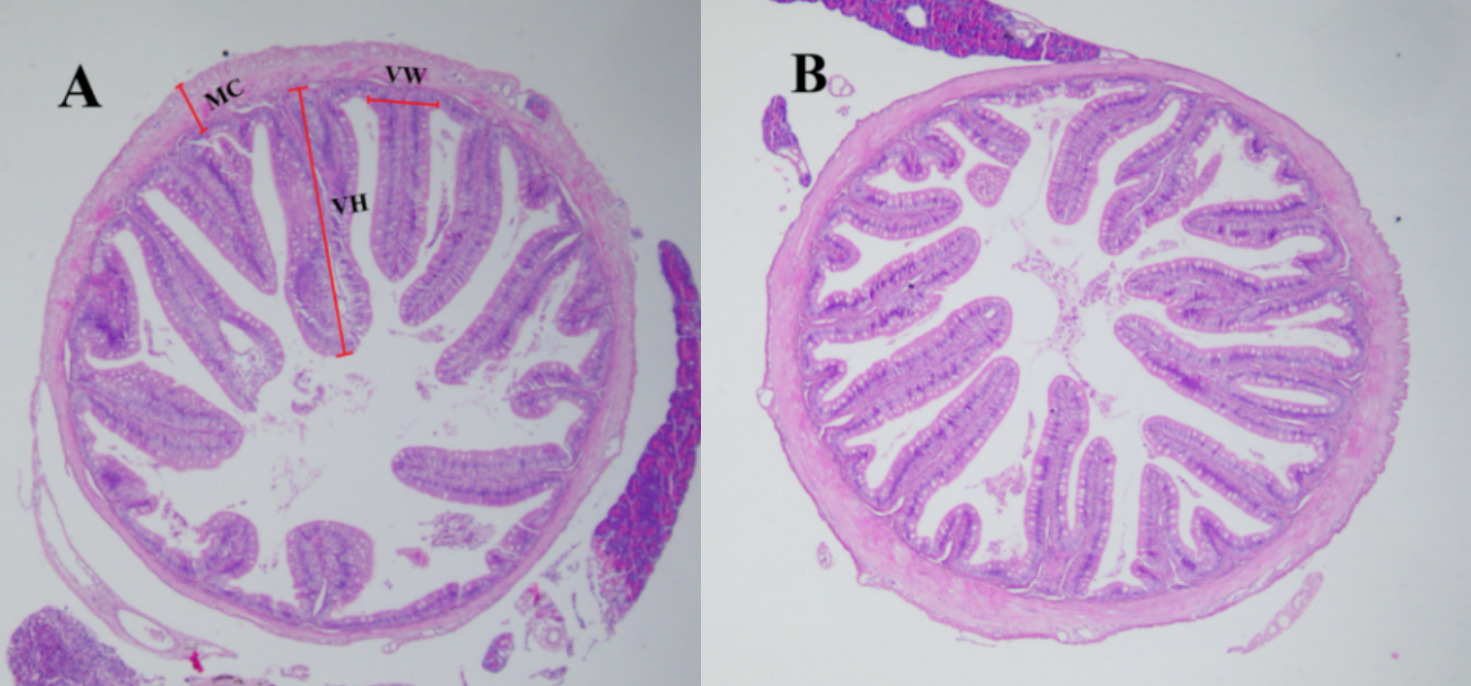
Three fish from each group were randomly selected for histological examination on day 60. The midgut tissues were dissected from each fish and fixed in 4% paraformaldehyde solution for 24 h, dehydrated through a graded ethanol series, cleared in xylene, and embedded in paraffin wax. Sections were cut at a thickness of 5 μm using a rotary microtome (Leica RM2235, Germany), stained with hematoxylin and eosin (H&E), and examined under a light microscope (Olympus, Japan). Image-Pro Plus 6.0 software was used to measure villus height (VH), villus width (VW), and muscularis thickness (MC) of the gut at 40× magnification. Then, three fish from each group were collected for microbial composition analysis on day 30 and day 60.
Sequencing was performed by Biomarker Technologies Co., Ltd. (Beijing, China). Total genomic DNA was extracted from intestinal contents using the TianGen DNA Kit (TianGen, China) following the manufacturer’s protocol. The V3-V4 hypervariable region of the bacterial 16S rRNA gene was amplified using the universal primers 338F (5’-ACTCCTACGGGAGGCAGCA-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’). PCR amplification was performed under the following conditions: initial denaturation at 98°C for 2 min, followed by 30 cycles of denaturation at 98°C for 15 s, annealing at 55°C for 30 s, and extension at 72°C for 30 s, with a final extension at 72°C for 5 min. Sequencing libraries were prepared and sequenced on the Illumina NovaSeq 6000 platform (Illumina, USA) using paired-end 250 bp reads. The raw data obtained from high-throughput sequencing were converted into sequencing reads through base-calling analysis. The raw data were then filtered, primarily by Trimmomatic0.33, according to single nucleotide quality. Primer sequence identification and removal were performed using Cutadapt1.9.1. PE reads generated in previous steps were assembled with USEARCH 10.0, and chimeras were removed with UCHIME 8.1. The high-quality reads generated from the above steps were used in the following analysis. Finally, the obtained high-quality reads were analyzed in the following aspects: OTU analysis, species annotation and taxonomic analysis, diversity analysis, association analysis, significant difference analysis, and functional prediction.
2.2.5. Challenge assays
After the feeding experiment, the turbots were temporarily held for 1 week, and then the challenge experiment was conducted. The pathogenic bacterium Aeromonas salmonicida BHAS-1 was kept in the laboratory. BHAS-1 was plated on BHI plates for 48 h, and the pure culture was rinsed with sterilized normal saline to make 1.62×108 CFU/mL bacterial suspension. In our previous study, we calculated 1.62×108 CFU/mL as the semilethal concentration (LC50) of BHAS-1 by the karbers method.15 Fifty juvenile turbots were randomly selected from the CK group and TC group, respectively, of which 100 μL BHAS-1 was injected into all fish in both groups by the intraperitoneal injection method. The two groups were fed with a basal diet, and the other feeding conditions are described in section 2.1.2. The number of deaths in each group was observed and recorded daily after infection. After the turbot stopped dying, three individuals from each group were randomly selected for determination of intestinal flora. The 16S rRNA sequencing method was described in section 2.2.4.
2.3. Statistical analyses
All statistics were analyzed by GraphPad Prism8.0.2. Statistical differences were assessed using t-test, and the level of statistical significance was set at P ≤ 0.05.
3. Results
3.1. Selection of potential probiotic bacteria
After a 30-day pre-experiment, the results showed that Halomonas titanicae (CMCC Name HT-Tc3, TC) significantly improved individual growth performance compared to other strains (Table 2). Therefore, TC was identified as the potential probiotic bacteria.
3.2. Evaluation of probiotic properties of HT-Tc3
3.2.1. Security evaluation
A hemolytic test was conducted on agar plates containing sheep blood, and no hemolytic zones were observed. This proves that the strain is harmless to human and can be used as a potential probiotic for further study.
3.2.2. Effect of HT-Tc3-supplementt on growth
On day 60, the growth performance of the fish was calculated. The TC group exhibited a 10.2% increase in length and a 34.8% increase in weight compared to the CK group (Table 3).
3.2.3. Effect of HT-Tc3-supplement on antioxidant activity
To analyze the effect of probiotic supplementation on antioxidant activity in fish, we determined the activities of SOD, CAT, and AKP (Fig. 1). On day 30, AKP activity showed a significant increase compared to the CK group (P < 0.05), while there was no significant change in SOD and CAT activity in the TC group (P > 0.05). On day 60, the TC group exhibited significantly higher SOD and AKP activity than the CK group (P < 0.05), whereas CAT activity showed a significant decrease compared to the CK group. (P < 0.05).

3.2.4. Effect of HT-Tc3-supplement on digestive enzyme activity
TPS, LPS and AMS were chosen to assess the effect of probiotics on intestinal digestibility (Fig. 2). On day 30, there was no significant change in AMS and TPS activity (P > 0.05), while LPS activity significantly decreased in the TC group (P < 0.05). On day 60, AMS and TPS activities showed a significant increase in the TC group (P < 0.05), whereas LPS activity showed no significant change (P > 0.05).

3.2.5. Effect of HT-Tc3-supplement on the structural morphology of the intestine
The results of histology in the intestine indicated that VH, VW and MC were increased in the TC group compared to the CK group (Fig. 3 and Table 4)
3.2.6. Effect of HT-Tc3-supplement on intestinal flora
All sequences were categorized into operational taxonomic units (OTUs) based on ≥ 97% similarity, with each OTU representing a unique unit of classification. Alpha diversity indices reflect species richness. The coverage index was ≥99.9% in all samples, indicating that microorganisms were adequately detected and that sequencing quality was good. The results showed that the number of OTUs and alpha diversity indices of TC group was higher than those of the CK group on day 30 and 60 (Table 5 and Table 6)
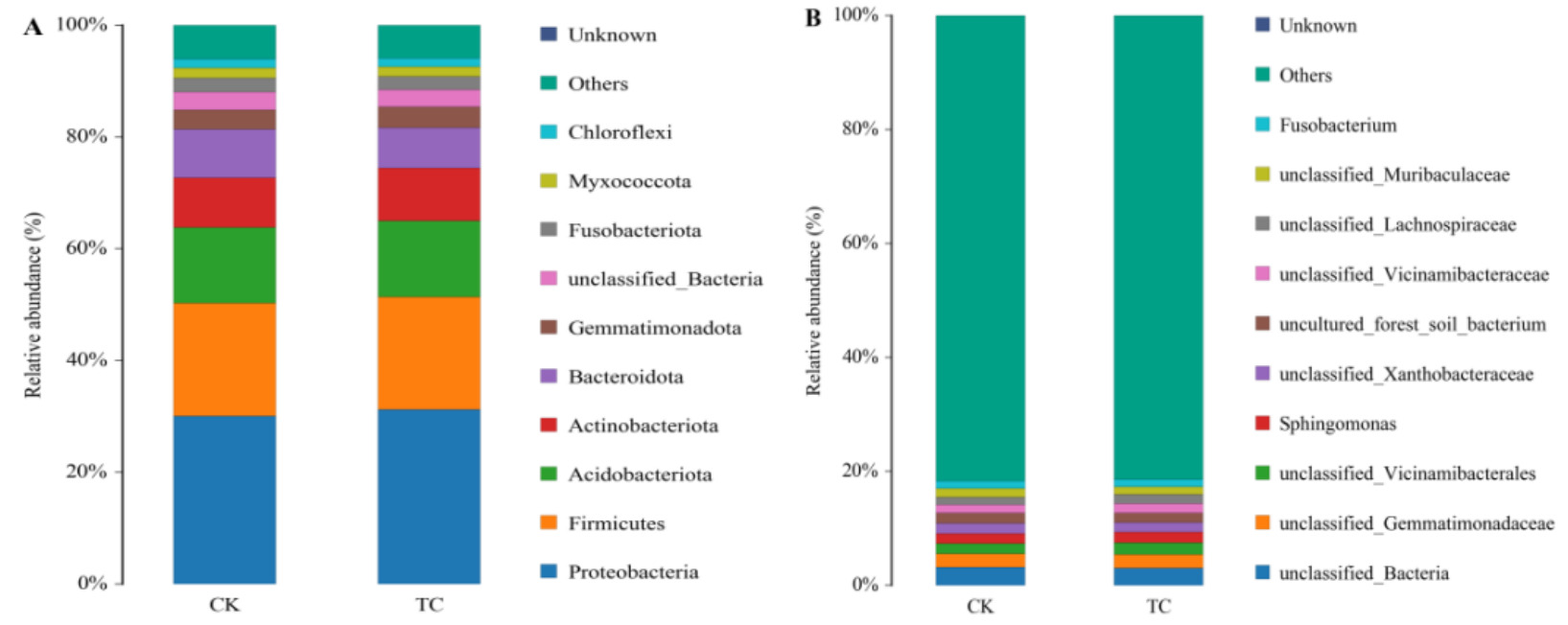
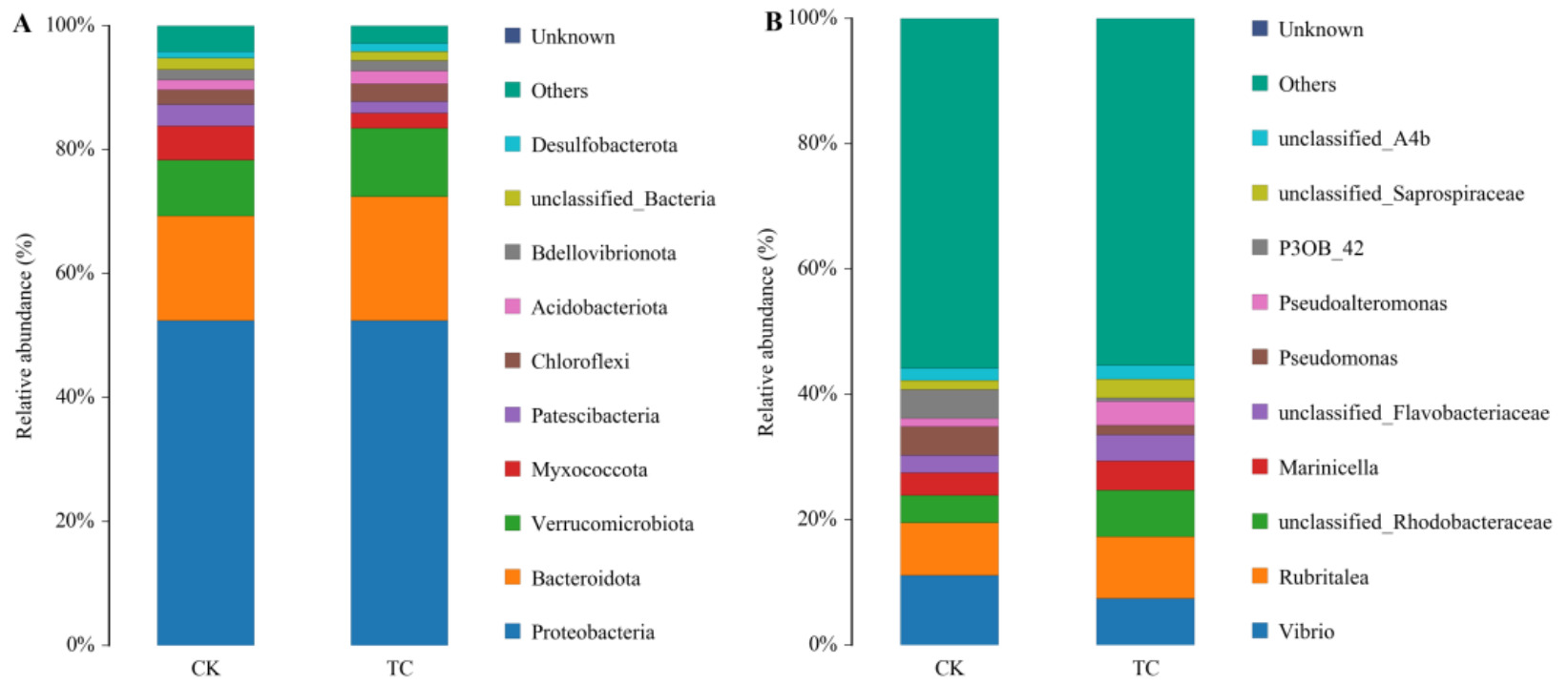
On day 30, at the phylum level, the dominant phyla Proteobacteria (31.25% vs 30.11%) and Actinobacteriota (9.45% vs 8.91%) exhibited higher relative abundances in the TC group than in the CK group. At the genus level, the relative abundances of unclassified_Vicinamibacterales (2.04% vs 1.85%), Sphingomonas (1.87% vs 1.72%, unclassified_Vicinamibacteraceae (1.68% vs 1.43%) and unclassified_Lachnospiraceae (1.58% vs 1.38%) were all higher in the TC group than in the CK group. The statistical distribution of the intestinal microbiota on day 30 is presented in Fig. 4. On day 60, there was a significant change in the structure of the intestinal microbiota in both groups compared with day 30. At the phylum level, the relative abundance of Bacteroidota and Cyanobacteria increased from day 30 to day 60 within both groups. At the genus level, unclassified_Cyanobacteriales, Cyanobium_PCC_6307, unclassified_Rhodobacteraceae, and unclassified_Comamonadaceae, and Lactobacillus became the dominant microbiota. Compared with the CK group, the relative abundance of dominant bacteria Proteobacteria (36.71% vs 34.68%) and Firmicutes (18.86% vs 16.25%) was higher in the TC group at the phylum level, and the relative abundance of beneficial bacteria Lactobacillus (2.38% vs 1.73%) was higher in the TC group at the genus level. The statistics on the distribution of the intestinal microbiota on day 60 are shown in Fig. 5.
_and_genus_level(b)_.png)
_and_genus_level(b)_.png)
3.2.7. Challenge assay
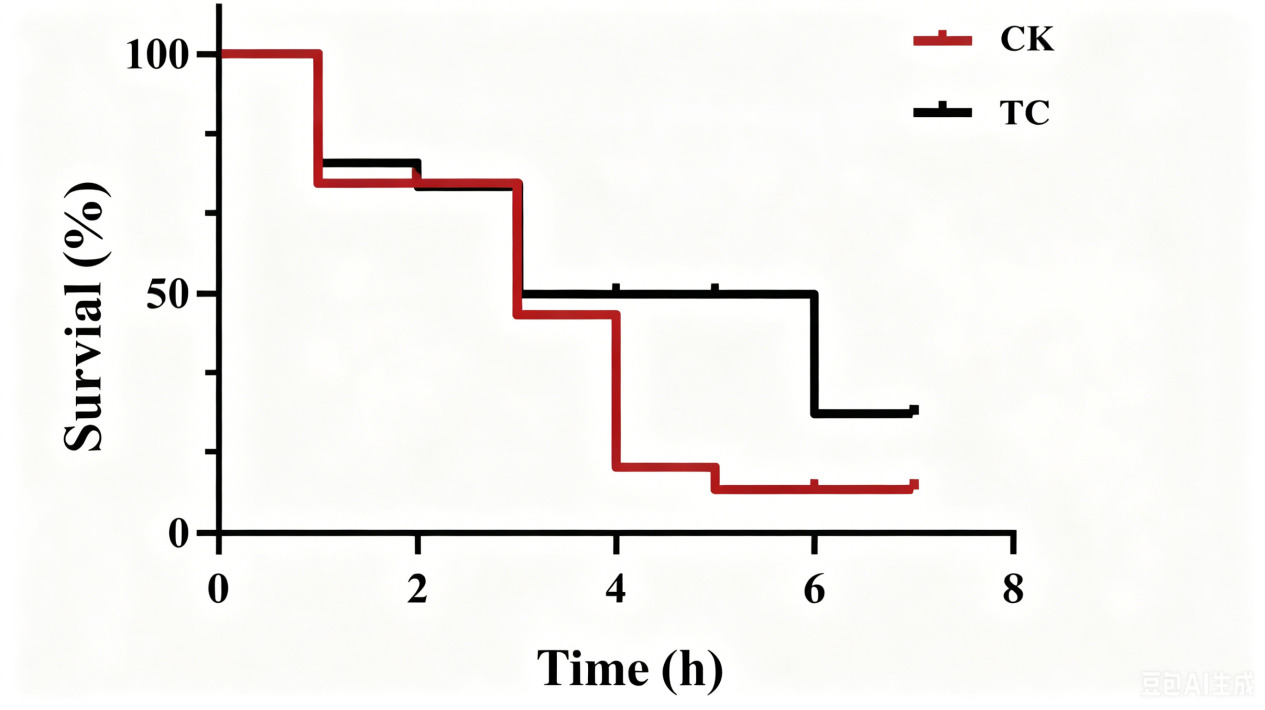
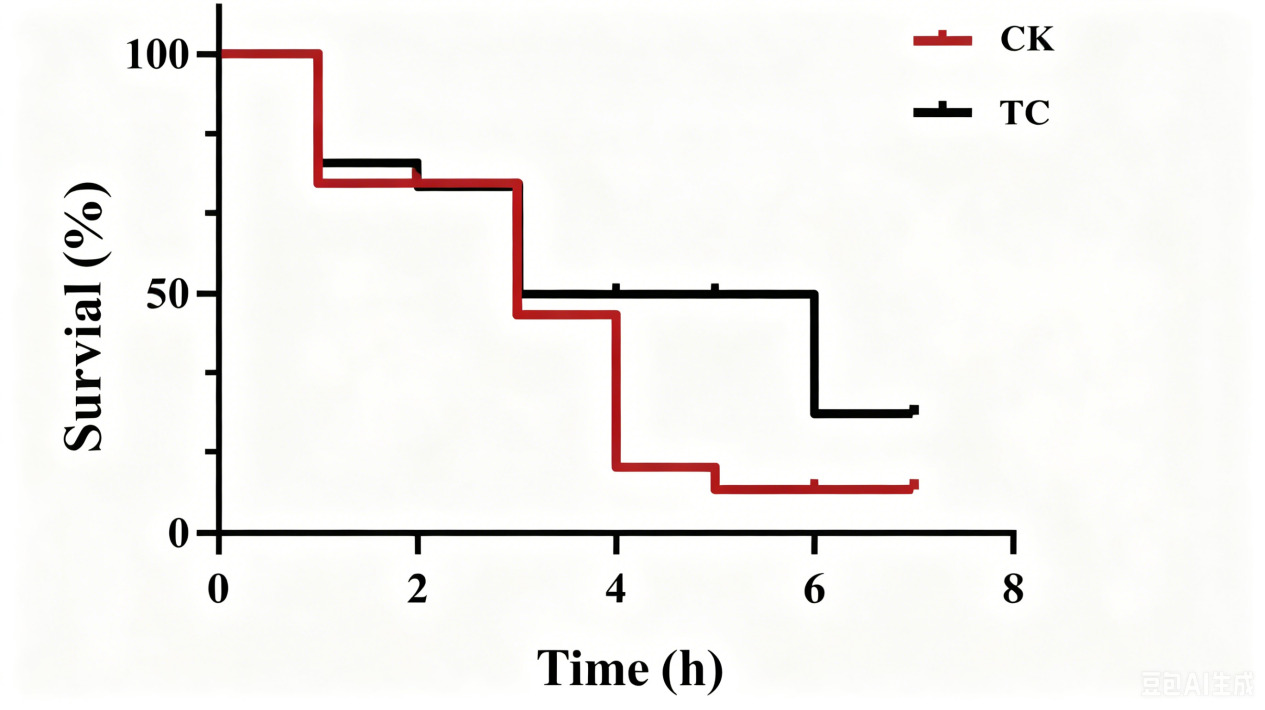
The TC group achieved a 16-percentage-point higher survival rate (28% vs. 12%) following challenge with A. salmonicida BHAS-1 (Fig. 6).
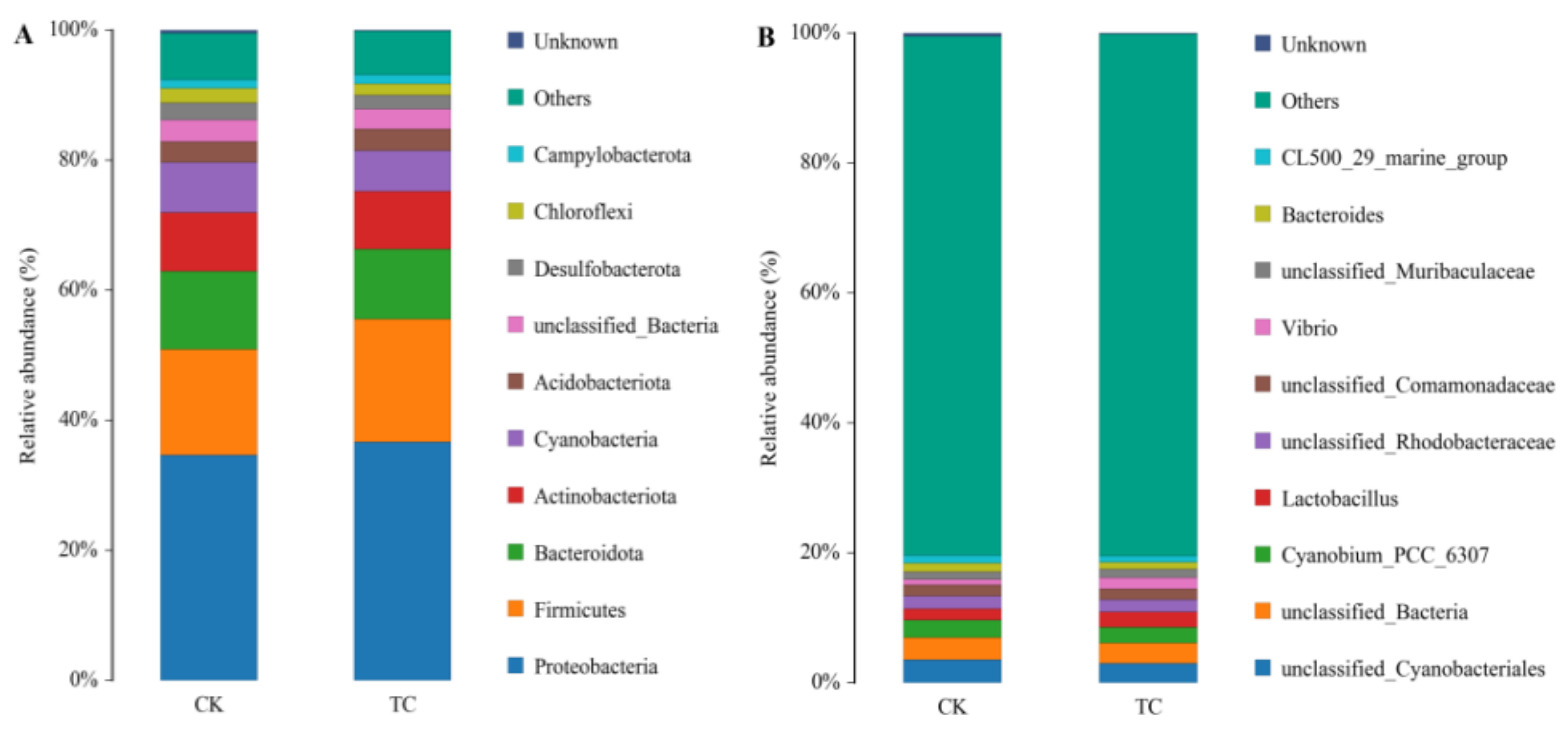
For the CK group and TC group, an average of 943 and 1,057 OTU numbers were obtained, respectively (Table 7), and the Alpha diversity indices of TC group was higher than those of the CK group (Table 8). The coverage index was ≥ 99.9% in all samples, indicating that microorganisms were adequately detected and the sequencing quality was good. At the phylum level, the relative abundance of the Bacteroidota and Verrucomicrobiota was higher in the TC group compared with the CK group. At the genus level, the dominant microflora differed between the two groups. In the CK group, the dominant genera were Vibrio (11.4%), Rubritalea (8.40%), Pseudomonas (4.62%), P3OB_42 (4.53%), and unclassified_Rhodobacteraceae (4.35%). The dominant genera in the TC group were Rubritalea(9.85%), unclassified_Rhodobacteraceae(7.45%), Vibrio (7.42%), Marinicella(4.67%), and unclassified Flavobacteriaceae (4.14%). The statistics of intestinal microbiota distribution are shown in Fig. 7.
_and_genus_level(b)_.png)
4. Discussion
The use of probiotics in the aquaculture industry has attracted interest, particularly for promoting environmentally friendly breeding practices. Halomonas titanicae, known for its ability to degrade organic pollutants and oil, is a promising probiotic.
A qualified probiotic strain must meet the following criteria: 1) It must be safe for humans. 2) It should be able to survive the intestinal tract of aquatic hosts. 3) It should provide benefit to the aquatic host.16 In this study, we conducted a comprehensive examination of H. titanicae HT-Tc3, laying the foundation for the development of microbiological products.
Probiotics are used as dietary supplements in aquaculture, and these aquatic products are eventually consumed by humans. Therefore, ensuring the safety of both humans and the host is of utmost importance in probiotic screening. Since HT-Tc3 was isolated from the turbot intestine, it can be confidently assumed to be harmless to the host. Our hemolytic tests have confirmed that this strain is safe for humans. No hemolytic zones were observed, which demonstrates that the strain is harmless to humans.
After confirming its safety, we evaluated the probiotic properties of HT-Tc3 from multiple aspects. Our feeding trial demonstrated that HT-Tc3 supplementation significantly improved growth performance, indicating its potential for development and application in aquaculture. Digestive enzymes, including proteases, amylases, and lipases, are critical for growth and serve as key indicators of feed utilization and digestive capacity. Previous studies have shown that probiotics can actively produce digestive enzymes such as amylase, pepsin, and lipase.17 Additionally, because digestive enzyme activities in fish can be modulated by dietary composition, probiotic supplementation may also stimulate the host to enhance endogenous enzyme production.18 On day 30, most digestive enzyme activities showed no significant differences between the TC and control groups, except for LPS activity, which was significantly inhibited in the TC group. Despite this inhibition, juvenile turbot in the TC group still exhibited robust growth. By day 60, however, AMS and TPS activities were significantly higher in the TC group than in the control group, and the suppression of LPS activity was no longer observed. The delayed enhancement of enzyme activities is likely attributable to the gradual process of intestinal adhesion and colonization. The adhesion and colonization of gut microbiota are dynamic processes that require a sufficient population size to achieve stable establishment.19,20 At the early stage of HT-Tc3 supplementation, the strain had not yet fully colonized the intestinal tract and thus could not fully exert its beneficial effects. Over time, however, HT-Tc3 became thoroughly established, the gut microbial structure stabilized, and positive effects on host digestion began to emerge.
SOD and CAT are essential components of the antioxidant system, which can contribute to resilience. Their main function is to eliminate harmful oxygen radicals produced during respiratory bursts, thereby protecting the structure and function of cell membranes from interference and damage caused by peroxides.21,22 Studies have suggested that the host’s commensal intestinal bacteria can stimulate the activation of defense mechanisms, and the extent of this activation depends on the bacterial species and its viability.23–25 On day 30 of our study, the expression patterns of antioxidant enzymes were similar to those of digestive enzymes. On day 60, SOD activity was significantly higher in the TC group compared to the control group. This trend may also be attributed to the mechanisms of gut flora adhesion and colonization. Interestingly, CAT activity was significantly lower in the TC group on day 60. This observation may reflect a compensatory regulation within the antioxidant defense network. Given the significant elevation of SOD activity, the efficient dismutation of superoxide radicals may have reduced the burden on the CAT pathway, a phenomenon previously documented in fish fed with probiotics.26 Furthermore, the immunoregulatory capacity of probiotics is known to vary with the level of supplementation.27 Therefore, the level of probiotic supplementation is selected to optimize its effects on improving production performance and preventing disease, which is also the main focus of our upcoming research. In addition to its role in antioxidant defense, AKP is a key enzyme involved in phosphorus metabolism and nonspecific immunity. In the present study, AKP activity was significantly higher in the TC group than in the control group on days 30 and 60, suggesting that HT-Tc3 supplementation may enhance non-specific immune function in turbot.
In summary, HT-Tc3 enhances digestive absorption capacity and promotes nutrient conversion by increasing digestive enzyme activity. It also enhances resistance to external pathogens by increasing the activity of immune enzymes, thereby affecting the host’s physiological state.
Studies have reported that probiotics promote growth by altering the intestinal environment and the structure of the intestinal epithelium.28,29 Our study found that supplementing the diet with HT-Tc3increased the villi height, width, and muscular thickness, which improves digestion and nutrient absorption and ultimately enhances turbot growth. Although the improvements in intestinal structure were not statistically significant, a positive trend was observed.
The gut microbiota plays a crucial role in nutrient metabolism, energy absorption, cholesterol metabolism, and in tracking and enhancing the host immune response.30–32 Therefore, studying the gut microbiota is essential for evaluating the effectiveness of probiotics. The results showed that the TC group had higher OTU counts and species diversity index than the CK group. This result suggested that continuous supplementation with HT-Tc3 significantly increased intestinal flora diversity and effectively enhanced the balance of microecological zones, resulting in a series of positive effects. On day 30, the phylum structure of the intestinal flora between the two groups was similar. The dominant phyla in both groups were Proteobacteria, Firmicutes, Acidobacteriota, Actinobacteriota, and Bacteroidota, which is consistent with previous studies on the intestinal flora of turbot.33 Our study also found that Acidobacteriota was among the dominant bacteria, a finding not reported in previous studies. The presence of Acidobacteriota may be related to our specific culture conditions. Members of this phylum have recently been recognized for their potential role in the degradation of complex polysaccharides and nitrogen cycling in aquatic environments, which may contribute to gut health and nutrient utilization.34 The reason for this may be related to our cultural patterns. Several external factors can affect the establishment and nature of the microbial composition, such as the surrounding environment, feed, and rearing water.35 In the TC group, the relative abundance of Proteobacteria and Actinobacteria was higher than that in the CK group. Proteobacteria and Actinobacteria have been shown to stimulate immune system development and maintain normal immune function.36–39 At the genus level, there was no difference in the composition of the dominant flora between both groups. HT-Tc3 dietary supplementation increased the relative abundance of unclassified_Vicinamibacterales and Sphingomonas compared to the CK group. Sphingomonas, as a new microbial resource for the biodegradation of aromatic compounds, occupies a more dominant position in the TC group. With their high metabolic capacity and versatile physiological properties, Sphingomonas has the potential to control and restore the mariculture environment.40–42 On day 60, there was a gradual change in gut flora structure compared to day 30, with Proteobacteria and Firmicutes remaining dominant, and a significant increase in Bacteroidota and Cyanobacteria. The number and composition of gut microbial communities change dynamically in response to individual host development. The continuous addition of HT-Tc3 further increased the abundance of Proteobacteria and Firmicute, which are part of the core microbiome. In the intestinal microbial system, the core microbiome plays a crucial role in controlling the entire bacterial community. If the core gut microbial communities disappear, the entire bacterial community structure may disintegrate.43 At the genus level, the gut microbiota also underwent succession as the host developed, with differences between the groups. The dominant bacteria in two groups were unclassified_Cyanobacteriales, Cyanobium_PCC_6307, unclassified_Rhodobacteraceae, and Lactobacillus. In the TC group, the abundance of Lactobacillus was significantly higher than that in the CK group. Lactobacillus is a widely used microorganism known to improve fish growth and disease resistance in commercial aquaculture. It is worth noting that Vibrio abundance increased in the TC group. Although Vibrio sp. isolates are commonly believed to be pathogenic to aquatic animals, the experimental fish were in good health, and no significant disease symptoms or deaths occurred during the experiment. Previous studies have also shown that Vibrio sp. make up a higher proportion of gut microorganisms than freshwater fish and often act as the dominant flora in marine fish, especially in turbot.44–46 Some non-virulent Vibro spp. are also believed to have probiotic properties.47,48 For example, the dietary supplementation of V. tasmaniensis HS10, isolated from the intestine of sea cucumber, can improve the innate immunity of sea cucumber.49 Additionally, a comparative metagenomics analysis of turbot revealed quorum-sensing and biofilm-formation system genes belonging to Vibrio sp.50 Therefore, we believe that Vibrio sp. plays a regular role and promotes various metabolic functions under stable aquatic conditions. When the environmental stability is disrupted, Vibrio sp. may be stimulated by these external stresses to express virulence genes through cascade reactions that ultimately lead to disease or death in turbot. This hypothesis needs to be explored in more depth. Nevertheless, monitoring and managing the breeding environment should remain of utmost importance.
It has been established that colonization and persistence in the gut are essential criteria for evaluating probiotics. If the strain does not effectively colonize the gut over time, it will be metabolized by the host and eliminated from the body. This situation has two negative effects. Firstly, probiotics need to be added frequently to ensure their effectiveness. Secondly, the frequent addition of probiotics increases farmers’ expenses. Additionally, some studies have highlighted an overlooked fact that abruptly stopping probiotic administration may increase susceptibility to host pathogens by causing gut dysbiosis, particularly in immunocompromised populations such as juvenile fish.51 Therefore, we conducted a challenge assay to assess colonization and disease resistance and to determine if discontinuing intake of the HT-Tc3 strain made the gut flora more vulnerable. It is worth noting that the TC group achieved better results in this test. The survival rate of the TC group was higher than that of the CK group after turbot were infected with A. salmonicida BHAS-1. 16S rRNA sequencing results showed higher OTU numbers and a higher species diversity index in the TC group than in the CK group. At the phylum level, the TC group had a higher relative abundance of Firmicutes and Bacteroidota than the CK group. Firmicutes, which are crucial for maintaining intestinal flora stability, play a decisive role, especially under external stress. Some members of the Bacteroidota have been shown to enhance specific immunity.49 At the genus level, the superiority of the TC group was even more evident. Many Pseudomonas species are also among the common pathogenic bacteria in aquaculture.52
The destruction of the immune barrier caused by the Aeromonas salmonicida BHAS-1 attack enabled pathogenic bacteria to enter, leading to an increase in Vibrio abundance in the CK group. Excitingly, supplementation with HT-Tc3 reduced the relative abundances of opportunistic pathogens Vibrio and Pseudomonas compared with the CK group, effectively halting disease progression. Other dominant genera, such as Rubritalea, unclassified_Rhodobacteraceae, Marinicella, and unclassified_Flavobacteriaceae, were also relatively abundant in the TC group compared to the CK group. Rhodobacteraceae and Flavobacteriaceae are known to enhance non-specific immune functions and resistance to infection.53 Although the roles of Rubritaceae and Marinicella in aquatic organisms are largely unknown, we hypothesize that these two genera may be closely associated with the health status of aquatic organisms. Thus, the challenge assay showed that HT-Tc3 can enhance resistance to pathogenic strains. This test also demonstrated the strain’s strong colonization ability and sustained beneficial effects, even after HT-Tc3 suspension.
5. Conclusions
In conclusion, this study provides the first evidence for the practical application of H. titanicae HT-Tc3 in turbot aquaculture. Dietary supplementation with HT-Tc3 for 60 days increased body length by 10.2% and improved weight gain by 34.8%, accompanied by significantly enhanced digestive enzyme activities. Improvements were also observed in intestinal morphology and the composition of the gut microbiota. Notably, the TC group achieved a 16-percentage-point higher survival rate (28% vs. 12%) following challenge with A. salmonicida BHAS-1, even after probiotic administration had been withdrawn. These findings demonstrate the long-lasting beneficial effects of H. titanicae HT-Tc3 and support its potential as a functional feed additive for sustainable marine aquaculture.
Acknowledgments
The authors gratefully acknowledge financial support from the Yantai Science and Technology Innovation Development Program Project (grant number: 2024YD079).
Authors’ Contribution - CRediT
Writing – original draft: Hanzhi Xu (Lead). Data curation: Dongliang Wang (Equal), Han Wang (Equal). Investigation: Xin Xu (Equal), Yujie Ding (Equal), Yanwen Zhuang (Equal), Yanan Cao (Equal). Visualization: Liwen Zhang (Equal), Fan Zhang (Equal). Resources: He Wang (Equal), LIdong Sun (Equal). Writing – review & editing: He Wang (Equal), LIdong Sun (Equal).
Competing of Interest – COPE
No competing interests were disclosed.
Ethical Conduct Approval – IACUC
This study was performed in accordance with the Guidelines for Care and Use of Laboratory Animals of the Chinese Association for Laboratory Animal Sciences (No. 2011-2).
Informed Consent Statement
All authors and institutions have confirmed this manuscript for publication.
Data Availability Statement
All are available upon reasonable request.
el(A) and genus level(B) after challenge assay