Introduction
The Yuanshui River, the second largest freshwater river in China’s Hunan Province, has a wide variety of fishery resources, acting as an essential germplasm repository for fish in the Yangtze River system.1 In recent years, the construction and operation of many cascade hydropower stations and large reservoirs have been built in the mainstream of the Yuanshui River, and the connectivity of the river has been blocked. They cause great changes in the ecological environment factors of the Yuanshui River, such as flow velocity, water temperature, and other hydrological conditions, which have seriously affected the life activities and reproduction of fish in the Yuan River basin.2 Due to the water environment’s unique physicochemical properties, the fish’s swimming movement reflects its state of life activity and history.3 Once the natural water temperature changes, most fish will adjust their physiological systems and biochemical reactions to adapt to the new environmental temperature,4 thus affecting the swimming ability of fish. The general evaluation parameters of fish swimming ability include induction velocity, preferred swimming speed, critical swimming speed, and maximum swimming speed, etc.,5–8 These parameters are certainly helpful to explore fish’s active adaptation to river environment under cascade hydropower development.
Siniperca knerii Garman is a genus of Perciformes, Serranida, Siniperca, commonly known as Baigui, Bigeye Mandarin fish, etc. It is an essential economic fish distributed in the middle and upper reaches of the Yangtze River system. It is also one of the typical representatives of fishery resources of Yuan River under the development of a hydropower cascade.9 Its biology has been extensively studied in recent years in China, mainly focusing on morphology, age, growth, physiology, resource status, phylogeny, etc.10–13 Few studies have comprehensively evaluated the swimming ability of juvenile fish Siniperca knerii Garman, and there are few reports on the status of the effect of environmental temperature changes on the swimming ability and metabolic function of wild Siniperca knerii Garman under cascade development. To investigate the effect of water temperature changes on the swimming ability of wild Siniperca knerii Garman under cascade development, the induction velocity, preferred velocity, maximum swimming speed, and oxygen consumption rate of wild fish at different water temperatures were measured using a self-circulating fish-locomotion testing water tank. It can provide basic information for the adaptive evolution of locomotor patterns of fish and data for the conservation of fishery resources in the Yuanshui River.
Materials and Methods
Experimental fish
347 juvenile Siniperca knerii Garman were collected in the middle reaches (Hefu region)of the Yuanshui River from May to August 2019. A total of 133 juvenile fishes with similar body length (9-13cm) and good activity was selected for the experiment, which was completed in the laboratory of the Collaborative Innovation Center of the Aquatic Efficient Health Production in Hunan province of China at the same period. The morphological parameters of the experimental materials are shown in Table 1.
The fishes were temporarily reared in two aquariums (194cm×80cm×62cm) at a constant temperature. To reduce the stress response of the fish, the captured wild fish were acclimated for 7d before starting the experiment. During the period, common commercial fish pellet feed was fed. The multifunctional dissolved oxygen analyzer (Swiss, Mettle rSG98-FK2) was used to regularly measure its water temperature, dissolved oxygen and pH, and other water quality parameters. Constant temperature (20.0±1.0°C), natural photoperiod, sufficient oxygen supply(≥6.5mg/L), and pH (7.3±0.1) were maintained.
Experimental design
Based on the annual average water temperature range of Yuanshui River, four experimental water temperature levels (15, 20, 25, and 30°C) were set. After 7 d of acclimatization, the fish were divided into 4 experimental groups of 32-36 fish, and the temperature was increased by thermostatic heating rod and decreased by ice mass. The water temperature was regulated by 1-2℃ every day until the set temperature and then adapted to 30 d. The water temperature of the water tank for the self-circulating fish-locomotion test was maintained at this temperature at the same time. Then they were transferred to the tanks, and the swimming parameters s were measured. Flow velocity was measured with a velocity meter (China, LS300-A), oxygen content in the water was measured with a dissolved oxygen analyzer (Swiss, MettlerSG98-FK2), and a computer-connected camera (Sony ILCE-6300) was installed above and to the side of the tank to monitor and test the animals from multiple angles.
Experimental device
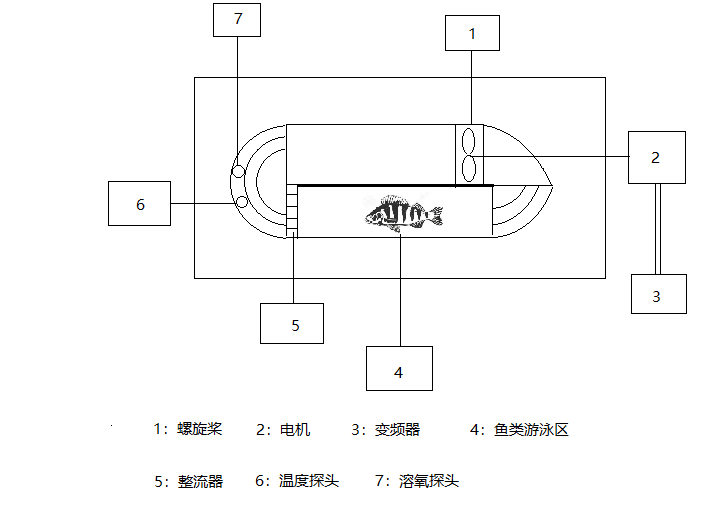
A self-circulating fish-locomotion testing water tank was used for this experiment (Figure 1). The device was a self-circulating hermetic made of stainless steel and acrylic transparent material. The propeller motor power was adjusted by an external frequency changer to change the rotational speed of the propeller, thus creating a circulating water flow with different flow velocities. And because of the rectifier, the flow will be relatively smooth and equal. The water flow velocity in the swimming area was calibrated using a flow meter and a frequency changer. The water flow velocity at different inverter frequencies was calculated using the conversion formula and the positive linear correlation between the motor frequency. The velocity was fitted: y=102.2171x-3.122 (R2=0.9903), and then the swimming velocity of the fish was determined. The dissolved oxygen analyzer monitored the changes in dissolved oxygen during fish swimming at different water flows. The volume of the sealed area of this device was 19.9L. The dimensions of the measured swimming area were 0.56m × 0.13m × 0.1m. In the experiment, the fish was placed in the fish observation area of the tank for 24h in standing water to eliminate the stress on the fish body during the transfer process, and then adapted to the flow rate of 0.5BL for 10min-30min, and then slowly increased the flow rate. The fish have enough time to adjust to the tank’s new environment to avoid stress, controlled remotely by a computer with cameras and free of human interference.
_propeller__2)_propeller_motor__3)_freque.png)
Swimming ability parameter induction velocity (Ui)
It also refers to the starting velocity. There are two parameters of induction velocity: absolute induction velocity (cm/s) and relative induction velocity (BL/s, BL is the length of the fish body). The “incremental velocity method” was mainly used.7 In the test, one fish was randomly selected and placed alone in the fish swimming area. After 24h adaptation, the fish kept heading toward the direction of water flow in the testing tank. Then the frequency changer gradually increased the frequency with a 0.1Hz gradient to change the water velocity in the fish swimming area until the fish adjusted its direction. When the reversing water flow (rheotaxis) time of the fish exceeds 10s, it is the induction velocity of the fish. the number of replicates was 12.
Preferred swimming speed (Uopt)
It is the most suitable water flow velocity for fish inhabitation and schooling. In preferred swimming speed, the fish can keep swimming in the direction of reversing water flow (rheotaxis) time for a long time. In this experiment, the standard measurement time is 30 min, and it is also divided into absolute (cm/s) and relatively preferred swimming speed (BL/s).
Specific method8: The juvenile fish was placed into the device first for 2h’s adaptation, and then adjust the frequency with a gradient of 0.2Hz, until the juvenile fish can continue swimming to reverse the water flow for 30min, the water velocity at this time can be regarded as absolute preferred swimming velocity. The relative preferred swimming speed was the absolute preferred swimming speed divided by the body length of the experimental fish. the number of replicates was the same as the induction velocity.
Critical swimming speed (Ucrit)
Specific method14: Before the formal experiment, each group’s critical swimming speed (Ue) was predicted. A healthy and uninjured juvenile fish was placed in the swimming area and adapted at a low velocity of less than 0.5BL/s for 2h. The velocity was increased by 0.4BL/s every 2 min until the fish became fatigued and stopped moving. The △U (15% Ue) during the formal experiment was determined accordingly.
When formal experiments were started, a juvenile fish was placed at a velocity of 0.5 BL/s for 30 min and then adjusted by 0.5 BL/s every 5 min, and 15% Ue was increased every 30 min after the 60% predicted value until the fish reached a state of exhaustion. The results were recorded, and the critical swimming speed (Ucrit) was calculated.
The calculation formula is as follows
Ucrit=Ui−1+(T/ΔT)×ΔU(1)Urcrit=Ucrit/BL(2)
Ucrit indicates absolute critical swimming speed, Ui-1 indicates the sub-high swimming speed of the fish before exhaustion. T indicates maximum swimming speed duration of the fish (less than 30 min), △T indicates 30 min, △U indicates 15% Ue predicted value, Urcrit indicates relative critical swimming speed, and BL indicates the fish body length.
The maximum swimming speed
It was measured by the incremental flow velocity method.6 A fish was placed in the swimming area and acclimated in a flow velocity of 0.5BL/s for 2h, the flow velocity was gradually adjusted to 1BL/s. In the end, the flow velocity was increased by 0.05 BL every 15s until the fish stopped swimming due to fatigue, and the flow velocity was regarded as the maximum swimming speed of the fish.
Oxygen consumption rate (OCR)
Specific method15: The oxygen consumption rate (30 min) was calculated by measuring the dissolved oxygen in the swimming area every 2 min, using different parameters such as V of the device water and the absolute value of the slope of dissolved oxygen with T (St) based on the formula; Subsequently, the device was run for 30 min without test fish at the corresponding flow velocity to exclude the microbial oxygen consumption.
The calculation formula for OCR (mg/kg.h)
OCR=(St×60−Sb)×V/M
“St” indicates the slope of dissolved oxygen within the swimming zone for each group (30 min) with T, (satisfying the correlation index R2 > 0.95). 60 indicates 60 min, and Sb indicates the slope of microbial oxygen consumption within the water tank. V indicates the volume within the swimming zone after removing fish volume, and M indicates fish quality (kg).
Tail beat frequency (TBF) and respiration frequency (RF)
Method: replay counting method.
The formula is as follows:
TBF( Hz)=TBT/TRF( Hz)=RRT/T
TBT is the tail beat frequency of a fish during the observation, and T is the time (s)
RRT is the respiration frequency of a fish at the observation time, T is the time (s)
Data processing
Data were analyzed by Excel 2010 and spss19.0. The data of each group were expressed by mean ± SD, and the differences between the data of each group were adopted one-way ANOVA and Duncan multiple comparisons. P<0.05 denotes significant differences and P>0.05 indicates insignificant differences.
Results
As shown in Table 2, the results showed that water temperature was negatively correlated with both absolute induction velocity and relative induction velocity of Siniperca knerii Garman in the temperature range of 15-30°C. They decreased with the increase in temperature. The relative induction velocities of Siniperca knerii Garman were (1.16±0.07), (0.94±0.06), (0.86±0.07), and (0.70±0.09) BL/ at the four temperatures, respectively. The parameters at 15°C were significantly higher than the other three groups and the lowest at 30°C. Preferred swimming speeds, critical swimming speeds, and maximum swimming speeds of the fish increased with increasing temperature (Table 2).
The relative preferred swimming speeds were (1.86±0.04), (2.23±0.35), (2.65±0.28) and (2.74±0.33) BL/s at four water temperatures, respectively. The relative critical swimming speeds of the fish were (2.58±0.32), (3.31±0.41), (4.31±0.52) and (5.22±0.48) BL/s for the four temperatures, respectively. The relative maximum swimming speeds at the four temperatures were (3.14 ± 0.22), (3.76 ± 0.26), (5.29±0.32) and (6.10±0.24) BL/s, respectively. The parameters of 15°C were all significantly lower than the other three temperatures (P<0.05), and the parameters of 30°C were the highest. As shown in Table 3, the regression models of other swimming parameters and water temperature were significant (P<0.05) except for induction velocity. The fitted equation for the relationship between water temperature(x) and absolute induction velocity(y) is y=0.0013x2-0.5023x+23.43 R2=0.4449, and the equation between water temperature and relative induction velocity is y=-0.0318x+1.8217 R2=0.3608. The fitted equation for the relationship between water temperature and absolute preferred swimming speed was y=-0.0677x2+4.4288x-40.026 R2 =0.7281, and the equation between water temperature and relative preferred swimming speed was y=-0.0075x2+0.4951x-5.0938 R2=0.775. The fitted equation between absolute critical swimming speed and the temperature was y=0.069x2-1.1239x+26.844 R2=0.9561. The equation between the relative critical swimming speed and the temperature was y=0.0047x2+0.0162x+0.4836 R2=0.9553. The fitted equation for the relationship between the absolute maximum swimming speed and the temperature was y=0.0029x2+0.1071x-0.2559 R2=0.9365; the fitted equation for the relationship between the relative maximum swimming speed and the temperature was y=0.0018x2 +0.0124x-0.0863 R2=0.912.
The oxygen consumption rate, respiration frequency, and tail beat frequency of Siniperca knerii Garman increased with increasing temperature. and the biggest value of the three index was found at 30°C, followed by 25°C,20°C, and 15°C. The oxygen consumption rate, respiration frequency, and tail beat frequency significantly differed among the four temperatures. The relationship between oxygen consumption rate(y) and critical swimming speed(x) in different temperature treatment groups is shown in Table 4. The equations of the relationship between oxygen consumption rate and relative critical swimming speed were y= 2.5391x2-148.91x+2649.1 for the 15°C group, y=-0.9751x2+91.097x-1516.1 for the 20°C group, y=0.1849x2-2.3545x+365.12 for the 25°C group, and y=0.3608x2-26.701x+1168.3 for the 30°C group.
The regression relationships between oxygen consumption rate (y)and relative critical swimming speed were all significant (P<0.05) for the four temperature groups. The relationship between oxygen consumption rate and tail beat frequency for different temperature treatment groups is shown in Table 5. y= 214.4x0.45 for the 15°C group, y=239.1x0.43 for the 20°C group, y= 379.32x0.39 for the 25°C treatment group, and y = 425.37x0.41 for the 30°C group. The regression equations for the four groups of oxygen consumption rate and tail beat frequency were all significantly correlated (P<0.05).
Discussion
Effect of water temperature on the induction velocity of Siniperca knerii Garman
As a result of the construction of the Yuanshui River water conservancy project, the river has been fragmented, and the habitat of local fish resources has been severely affected. The temperature stratification phenomenon is evident in the downstream reservoir area, such as Wuqiang Creek Reservoir, formed by its high dam. If the release of water is located below the “thermocline,” the water released in summer and autumn will be significantly lower than the natural water temperature. In contrast, winter and spring are the opposite. The water temperature is higher. Changes in water temperature in the reservoir area and downstream river sections have altered fish survival and reproductive conditions.1,2 Water temperature is one of the critical environmental factors affecting fish survival, distribution, and swimming ability.4,16,17 Siniperca knerii Garman is a genus of Perciformes and an important fish resource component of the Yangtze River and its affiliated river system. It often inhabits the deeper waters of the rivers in slow-flowing water or still-clean water as a benthic-ambush type of carnivorous fish with high basal and active metabolic levels.9 Meanwhile, in terms of ecology and predation, Siniperca knerii Garman prefers a flowing-water environment of rich dissolved oxygen level and has the habit of swimming against the current, often hunting for food by raiding and seizing the best spatial position of the habitat.12,15 The induction velocity, which is the flow velocity threshold of most rheotaxis fish, determines the minimum flow velocity of its rheotaxis and provides a basic reference for the measurement of fish Ucrit and Uopt.8 In this study, it was found that both absolute induction velocity and relative induction velocity of Siniperca knerii Garman decreased with increasing temperature. The data of 20°C were significantly higher than the other three temperatures. The data of 30°C were the lowest. The reason for the difference may be related to the temperature perception by the lateral line system of the fish, which relies on the lateral line system to perceive changes in the hydrodynamic characteristics of the habitat18 The results indicated that the temperature was higher and the high temperature la flow velocity could stimulate its initiative swimming. In contrast, low temperature inhibited the rheotaxis behavior of fish.
Effect of water temperature on the preferred swimming speed, critical swimming speed, and maximum velocity of the Siniperca knerii Garman
Water temperature also has a significant effect on other swimming parameters of fish. Studies have shown that fish critical swimming speed (Ucrit) and other parameters usually had a “bell-shaped” or a linear trend with temperature.19 “Linear-shaped” means that the Ucrit of fish can increase with water temperature. In contrast, “bell-shaped” means that the Ucrit of fish first increases with the increase of water temperature, reaches the maximum value, and then decreases. Preferred swimming speed is also called optimal swimming speed (Uopt). It refers to the swimming speed of fish moving forward in the most suitable water flow, which can determine their habitat choice to a certain extent. the flow velocity in the preferred habitat. The higher the swimming ability of many species of Cypriniformes, the better because most fish in the natural environment tend to swim at speeds close to Uopt during their search for bait and long migrations.20,21 In the optimal swimming speed range of fish, the oxygen consumption rate at a unit distance is minimal, and fish can swim continuously for a long time (≦200min).11 The critical swimming speed is mostly used as an indicator to evaluate the maximum sustained aerobic swimming ability and metabolism (red muscle) of fish and the relationship between environmental factors and the swimming ability of fish.4,5 The maximum swimming speed is mainly used to reflect and evaluate fish’s high-energy explosive anaerobic (white muscle) swimming ability, measuring the comprehensive performance of fish body locomotion and nervous system and the predatory and predatory anti-predatory ability of fish.6 Maximum swimming speed can be measured directly. Critical swimming speed can be calculated with the maximum swimming speed as the reference and the swimming time, speed increment, and other factors through the corresponding formula. The maximum swimming speed is the highest, the critical swimming speed is the second, and the preferred swimming speed is the least of the three speeds of fish.22 Water temperature is an important component of the environmental factors that affect swimming parameters. Most studies suggest that the critical swimming speed of fish generally occurs at water temperatures above 20°C. For example, Carassius auratus,14 Silurus meridionalis,14 and Salvelinus leucomaenis,4 From the results of this study it could, be seen that the critical swimming speed, preferred swimming speed and maximum swimming speed of Siniperca knerii Garman juvenile fish showed an approximately linear increasing trend with temperature. The data of 15°C was significantly lower than the other three temperatures (P<0.05), which was similar to most studies. For tropic fish, the maximum swimming capacity occurs mostly in the optimal water temperature range (25-30°C); for warm-temperate fish, it is between 20-30°C, and for cold-water fish, it occurs at 15-20°C22,23) but the influence of water temperature on the maximum swimming capacity of different species of fish varies greatly. At a certain water temperature, the maximum swimming index of most fish tends to decrease with the decrease in domestication temperature. Some research showed that low temperature has a significant reduction effect on the maximum swimming speed and critical swimming speed of juvenile fish of Myoxocephalus scorpius,24 Parabramis pekinensis20 and Spinibarbus sinensis,21 with a consequent reduction in the success rate of prey capture. The critical swimming speed of both Takifugu rubripes6 and Silurus meridionalis25 at different aquaculture temperatures was the lowest at 15°C and the highest at 30°C, similar to the present study. The usual explanation was that seasonal shifts in temperature cause significant changes in the quantity, distribution, and species of natural water bait from long-term temperature changes, and such changes often cause more conservative adaptive changes in the physiology and behavior of fish. Low-temperature limits fish’s energy activation and utilization, reduces the energy conversion rate and metabolism, and slows muscle activity. Low temperature also increases the water’s viscosity and the fish’s resistance to moving forward to reduce swimming ability. With the increase in water temperature, the energy conversion rate and energy concentration of fish gradually increase, the skeletal muscle contraction accelerates, and the swimming ability increases. In addition, most fish have a stronger predatory ability, higher metabolic capacity, and faster growth at higher summer temperatures in natural environments. The above physiological and biochemical regulation related to the domestication water temperature may be one of the important reasons why the speed parameters of each fish in the low-temperature group were smaller than that in the high-temperature group in this experiment. However, several scholars found that the critical swimming speed of Gambusia affinis and Lepomis macrochirus did not vary significantly in the domestication water temperature range of 18°C-30°C, which should be related to the species and domestication time.19
Effects of water temperature on oxygen consumption rate (OCR), tail beat frequency (TBF)and respiratory frequency (RF)of Siniperca knerii Garman
Water temperature is a key factor affecting fish’s physiological activities and ecological processes.26 The metabolic rate of most fish can increase significantly with the increase of water temperature in the suitable water temperature range.10,25,27 It is found that the oxygen consumption rate of Siniperca knerii Garman increased by 1.1-1.8 times during the increase of water temperature from 15°C to 20-30°C, respectively.11 TBF and swimming speed positively correlated in several teleostean fishes.28,29 This study found the same significant trend relationship in Siniperca knerii Garman. One reason can be that the change in water temperature leads to changes in the amount of dissolved oxygen in a specific temperature range. On the other hand, it can affect the mobilization of energy substrate supply and anaerobic metabolism time, affecting fish respiration metabolism and blood circulation to accelerate respiration frequency. With the increase of water temperature, the activity of various enzymes in the fish body is enhanced, and the metabolism is increased, which makes the tissue and muscle’s metabolic efficiency stronger. Therefore, the tail beat frequency is increased, the locomotion indicator is strengthened, and the energy demand to maintain fish activity and the balance of the body is increased, which leads to an increased oxygen consumption rate and an increase in the respiratory rate of fish. In this experiment, only the single environmental factor of water temperature in Yuanshui River water was considered, while other factors have corresponding effects on the role of fish swimming parameters. The next study should increase the experimental requirements of dissolved oxygen, transparency, flow velocity, water pressure, and other comprehensive conditions to obtain relevant basic data, which can be used to protect its resources effectively.
Acknowledgments
This work was supported by Foundation Project: National Natural Science Foundation of China (Grant No.31572619); Natural Science Foundation of Hunan Province (Grant No.2020JJ4446)
Conflict of interest
The authors declare no conflict of interest.