

Introduction
In fish classification, the difference in external morphology is the most intuitive. External morphology analysis is the simplest and most commonly used among the several fish population identification methods. Therefore, morphometry is often used to identify species and inter-species by measuring the phenotypic traits of the fish body. Morphological analysis frequently identifies inter-specific differences between different geographic populations, such as serranid fishes. Cavalcanti et al.1 determined the morphological traits of six serranid fish families, Ruiz-Campos reported morphological differences between different populations of Oncorhynchus mykiss,2 and Elliott performed morphological analysis on Orange roughy (Hoplostethus-atlanticus) captured from different geographic populations and found significant differences between them.3
Traditionally, morphometric methods were based on the method proposed by Hubbs et al.,4 measuring indices such as head length, body length, body height, and caudal peduncle from different directions and then comparing quantifiable and measurable traits individually. This single morphometric method is effective for species-level identification but often fails to identify subspecies-level populations.5 Multivariate statistical methods can compensate for this single identification method. Principal Component Analysis, Discriminant Analysis, and Cluster Analysis can more intuitively display the degree of differentiation and proximity between different populations. Neves used external morphometrics combined with principal component analysis and discriminant analysis to study geographical populations of Spondyliosoma cantarus captured in seven different geographical regions along the east-west Atlantic coast, showing that there were stable morphological differences among the four populations.6
Trachinotus ovatus, also known as the golden pompano, is widely distributed in the world, mainly distributed in the East and South China Seas in China. Its meat is tender and fat, grows rapidly, and is highly adaptable, making it one of the main economic fish species in Chinese marine aquaculture.7,8 During the growth and development of fish, external morphological characteristics are obtained through inheritance, which is mainly determined by genes. Studies have shown that differences in morphological characteristics are not entirely caused by heredity factors,5 and that environmental factors may also cause differences in morphological characteristics at key stages of fish growth and development.9,10 Therefore, because of the geographical distance and growth environment, the morphological characteristics of golden pompano populations in different locations may show a certain degree of differentiation. Still, research has yet to examine the morphological differences between different populations of golden pompano. Therefore, this study used traditional morphometric measurement methods combined with multivariate statistical methods such as principal component analysis, discriminant analysis, cluster analysis, and One-way ANOVA to compare the morphological differences between golden pompano populations in several coastal areas of China to provide a reference basis for the rational utilization of spatial population structure and population resources of different populations of golden pompano in coastal areas of China.
Materials and Methods
Sample collection
Golden pompano samples were collected from seven different geographic locations in China, including Zhanjiang City, Guangdong Province, Shenzhen City, Guangdong Province, Lingshui Li Autonomous County, Hainan Province, and Beihai City, Guangxi Zhuang Autonomous Region. Twenty 210 fresh samples of golden pompano were randomly selected from each sampling site, frozen, and shipped to the laboratory. Morphological characteristics were measured after thawing in the laboratory. The specific sampling times, locations, and body length information are listed in Table 1.
Data measurement
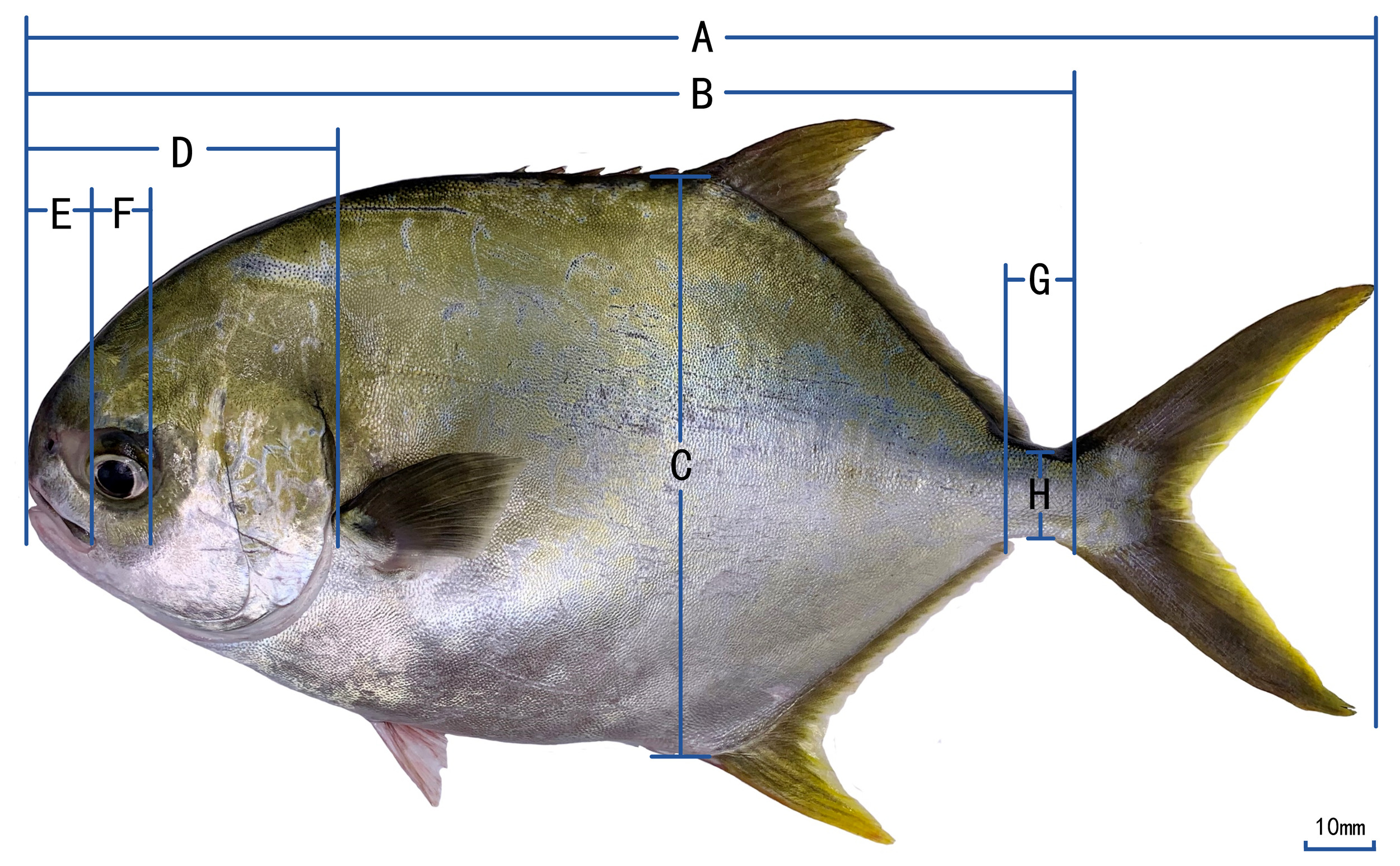
Traditional morphological methods are used to measure the golden pompano. The traditional morphological indicators measured include total length (TL), body length (BL), body depth (BD), head length (HL), snout length (SL), eye diameter (ED), eye spacing (ES), length of caudal peduncle (CPL), and depth of caudal peduncle (CPD). To make the measured data more accurate, the shape of the fish was straightened, and a ruler was placed under the fish during measurement. Images of fish and rulers were taken using a high-definition digital camera.11 The taken photos are imported into Adobe Photoshop 2021, and the length value of each trait parameter was calculated by using the number of pixels occupied by the scale of a ruler in the photo, which is accurate to 0.1 mm; an electronic digital vernier caliper measured the eye spacing, accurate to 0.1 mm. A schematic diagram of morphological measurements is shown in Figure 1.
Statistical Analysis
Microsoft Excel software was used to process the data to eliminate the influence of fish body size on character parameters during data analysis and improve data accuracy. The body length, body depth, head length, snout length, eye diameter, eye spacing, length of caudal peduncle, and depth of caudal peduncle were standardized by dividing the total length,12 and eight new proportional characters were obtained. The data of these eight proportional traits were imported into SPSS 24.0 software for multivariate statistical analysis, including principal component analysis, discriminant analysis, cluster analysis, and one-way analysis of variance (One-way ANOVA).
In SPSS software, “analysis-dimension reduction-factor” was used to conduct principal component analysis, and extract eight principal components and their eigenvalues, the contribution rate of each principal component, and the cumulative contribution rate according to the order of size. Scatter plots were drawn based on the scores of the first and second principal components to compare the variability and similarity of the seven golden pompano populations and to determine their morphological differentiation.13
Discriminant analysis of 210 samples from seven golden pompano populations was carried out using the stepwise discriminant method. The first two typical discriminant functions were extracted according to the eigenvalues, and scatter plots were drawn using these two functions.14 We established a discriminant classification function for eight proportional traits and predicted and classified the original samples according to the classification discriminant function. We calculated the discrimination accuracy rates P1 and P2 and the comprehensive discrimination rate.15 The calculation formula is as follows.
Discrimination accuracy P1:
P1=(AiBi)×100%
Discrimination accuracy P2:
P2=(AiCi)×100%
Comprehensive discrimination rate:
P=(n∑i=1Ai/n∑i=1Bi) × 100%
Where Ai is the number of tails correctly discriminated by the ith population, Bi is the number of tails involved in discriminating the ith population, Ci is the number of tails awarded to the ith population, and n is the number of populations.
The Euclidean distance was calculated using the shortest system clustering method of Euclidean distance among the seven populations. A population clustering relationship tree was constructed to compare the distance between the populations.
One-way ANOVA was performed using SPSS 24.0. To test the significant difference of each proportional character between different populations of golden pompano,16 0.05 was selected as the significance level, and 0.01 was selected as the highly significant level. Duncan’s method was used for multiple comparisons.
Results
Principal component analysis
In SPSS, through principal component analysis of eight proportional traits of golden pompano, eight principal components were extracted, and the cumulative contribution rate was 100%. The characteristic values, contribution rates, and cumulative contribution rates of each principal component are listed in Table 2. After sorting by the contribution rate of each principal component, the contribution rates of the first five principal components were 35.948, 22.244, 11.678, 8.949, and 6.971%, respectively. The contribution rates of principal components 4 and 5 were relatively small, indicating that the two principal components explain the morphological differences between different groups at low rates. The cumulative contribution rate of the first five principal components was 85.79%, which meets the requirement that the cumulative contribution rate is greater than or equal to 85%, indicating that the first five principal components can better explain 85.79% of the morphological differences between different golden pompano populations.
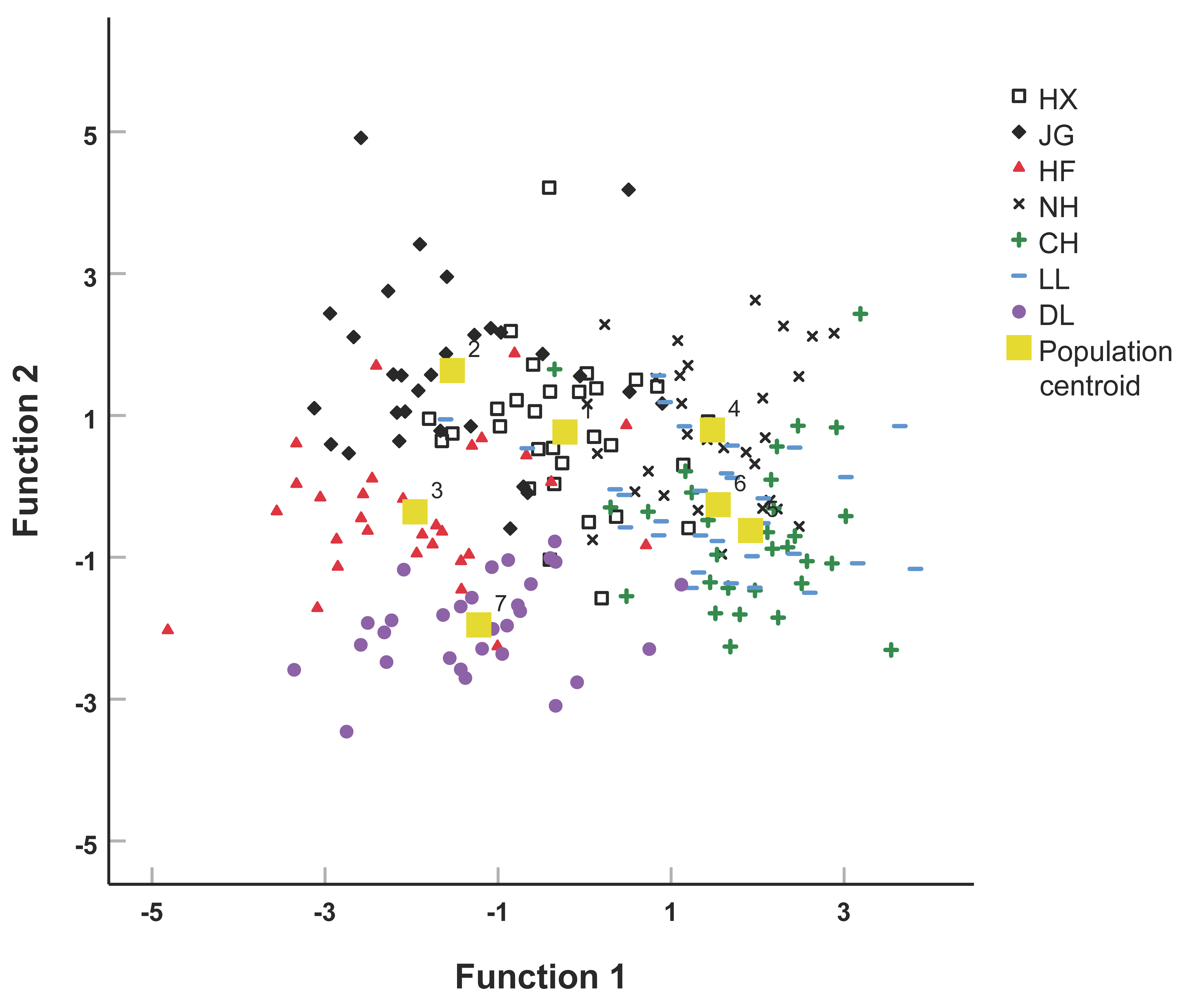
According to the factor loading values of each principal component, BL/TL, BD/TL, ED/TL, ES/TL, and CPD/TL, which play a key role in principal component 1, have corresponding loading values of 0.687, 0.670, 0.618, 0.694, and 0.646, respectively, all of which are greater than 0.600, and the contribution rate is relatively large. In other words, principal component 1 mainly explains the five proportional traits of BL/TL, BD/TL, ED/TL, ES/TL, and CPD/TL. The indicator with the highest contribution rate in principal component 2 was SL/TL, with a load value of 0.778. Principal component 2 mainly reflected the morphological characteristics of the snout. In principal component 3, the indicator with the highest contribution rate was CPL/TL, with a load value of 0.778, and principal component 3 mainly reflected the morphological characteristics of the tail. Therefore, the first three principal components could explain the seven proportional traits better. According to the factor scores of principal components 1 and 2, a scatter plot (Figure 2) can be drawn, intuitively showing the differences between different populations. According to the scatter plot of principal component 1 (PC1) and principal component 2 (PC2), there was a lot of overlap between the NH, CH, and LL populations, and the boundary between populations was not obvious, indicating that their morphological differences were similar. There was no overlap between the HX and LL populations, HX and DL populations, JG and LL populations, and LL and DL populations, indicating certain morphological differences between the above populations.

Discriminant analysis
Based on eight morphological proportionality traits, discriminant analysis was conducted on seven populations of golden pompano, and the stepwise discriminant method was adopted. The analysis results showed that all eight proportionality traits had significant meaning for the discriminant classification (P<0.05); therefore, all eight proportionality traits entered the discriminant function, and the discriminant classification function of golden pompano populations was constructed. According to these functions, the original samples were discriminated against and grouped. The discriminant function is expressed as follows:
HX: Y1=3759.706X1+744.047X2+262.180X3−1061.377X4−597.739X5+164.938X6−910.182X7−511.127X8−1597.083
JG: Y2=3759.960X1+738.779X2+143.813X3−783.050X4−587.596X5+418.094X6−1043.722X7−603.631X8−1583.675
HF: Y3=3774.060X1+708.990X2+246.614X3−1041.892X4−662.392X5+148.314X6−1022.173X7−796.591X8−1558.212
NH: Y4=3797.095X1+819.756X2+255.639X3−1146.883X4−659.821X5+16.440X6−894.243X7−391.353X8−1648.180
CH: Y5=3828.959X1+849.481X2+308.958X3−1346.703X4−735.093X5−258.601X6−972.328X7−436.528X8−1651.570
LL: Y6=3865.744X1+783.579X2+220.574X3−1239.062X4−744.746X5−68.427X6−905.849X7−385.145X8−1665.657
DL: Y7=3762.603X1+686.517X2+311.284X3−1167.400X4−875.665X5−73.284X6−997.412X7−557.578X8−1537.151
Where X1 = BL/TL, X2 = BD/TL, X3 = HL/TL, X4 = SL/TL, X5 = ED/TL, X6 = ES/TL, X7 = CPL/TL, and X8 = CPD/TL.
When determining the population affiliation of a sample, the numerical values of the eight proportional traits of the sample were substituted into the above seven functions, and the function with the maximum function value was the population category of the sample. The discrimination results are presented in Table 3. The comprehensive discrimination rate of the seven populations was 71.4%, and the discrimination accuracy rate of P1 was between 40% and 93.3%, while P2 was between 60.6% and 87.5%. Among them, the discrimination accuracy rates P1 of the JG, CH, and DL populations are all above 80%, and the discrimination accuracy rate P1 of the DL population is as high as 93.3%, and P2 is 87.5%; thus, the discrimination accuracy rate of the DL population is the highest. After discriminant analysis, the number of tails classified into each population was as follows: HX had 33 tails, accounting for 15.7% of the total; JG had 32 tails, accounting for 15.2% of the total; HF had 24 tails, accounting for 11.4% of the total; NH had 19 tails, accounting for 9.0% of the total; CH had 33 tails, accounting for 15.7% of the total; LL had 37 tails, accounting for 17.6% of the total; DL had 32 tails, accounting for 15.2% of the total.
In the stepwise discriminant analysis process of SPSS, two typical discriminant functions were extracted according to an eigenvalue greater than 1. The scatter diagram drawn according to these two typical discriminant functions is shown in Figure 3. It can be seen from the figure that the sample points of the populations of NH, CH, and LL are closely distributed and overlapping, and there is no overlap between the HX and DL population, JG population and the LL population, LL population and the DL population. This is consistent with the results of the principal component analysis, indicating that the NH, CH, and LL populations are closely related, and there are specific morphological differences between the other three populations.

Cluster analysis
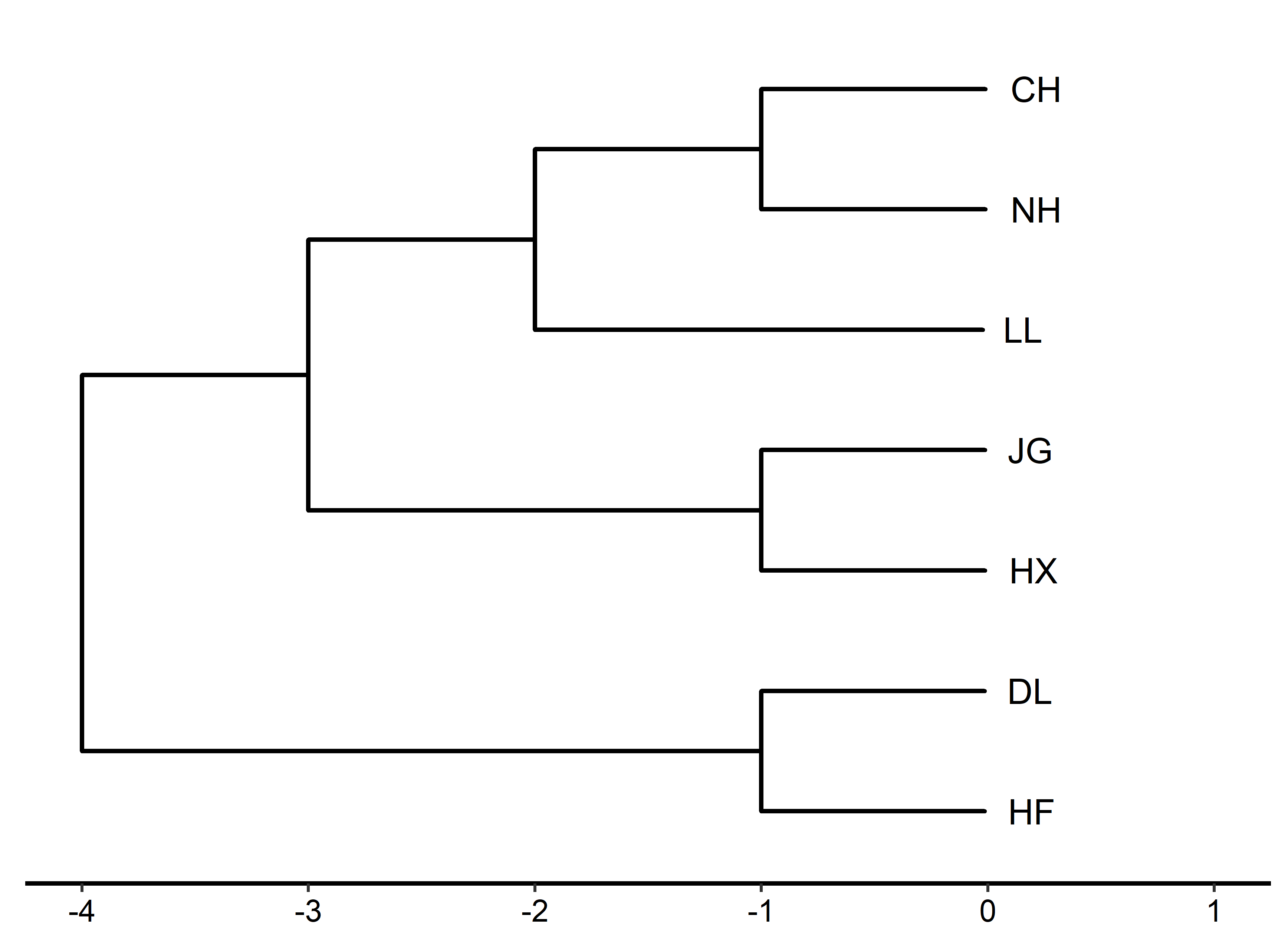
Cluster analysis can intuitively display the distance of morphological relationships between populations. This study performed cluster analysis on eight proportional traits of seven golden pompano populations to obtain the Euclidean distance between populations (Table 4). A dendrogram of the clustering relationship was drawn according to the Euclidean distance (Figure 4). It can be seen from Figure 4 that the CH and NH populations first formed a branch and then formed a branch with the LL population. JG and HX populations gathered into a branch, and so did DL and HF populations. The three large population branches were grouped. Overall, the seven golden pompano populations were divided into three groups: CH, NH, and LL, JG and HX, and LL and HF. The degree of genetic differentiation between the CH and NH populations, the JG and HX populations, and the DL and HF populations is the closest, and the genetic relationship is relatively close. The degree of genetic differentiation among the CH, NH, and LL populations is relatively close. The morphological difference is small, consistent with the principal component and discriminant analysis results.

One-way analysis of variance
Using the one-way ANOVA method in SPSS software, a single-factor variance analysis of eight proportional traits of seven golden pompano populations was conducted, and multiple comparisons of the proportional traits of each population were conducted. Results are shown in Table 5. The variance analysis results showed statistically significant differences (P<0.05) in all population proportion traits. Results of multiple comparisons showed differences in six indicators between the HX and LL populations, JG and LL populations, LL and DL populations, and JG and NH populations. The differences between the HX and LL populations were reflected in the BL/TL, HL/TL, SL/TL, ED/TL, ES/TL, and CPD/TL. Differences between the JG and LL populations were reflected in the BL/TL, SL/TL, ED/TL, ES/TL, CPL/TL, and CPD/TL. Differences between the LL and DL populations were reflected in the BL/TL, BD/TL, ED/TL, ES/TL, CPL/TL, and CPD/TL. Differences between the JG and NH populations were reflected in BL/TL, BD/TL, SL/TL, ES/TL, CPL/TL, and CPD/TL. Differences between the HX and DL populations were evident for all eight indicators, indicating that the morphological differences between these two populations were significant.
a,b,c,d,e,f In the same row, values with the same superscript mean no significant difference (P>0.05). In contrast, values with different superscripts mean statistically significant differences (P<0.05).
Discussion
The golden pompano is widely cultivated in southern China as a main economic fish species in Chinese seawater aquaculture. The main farming areas are Guangdong Province, Guangxi Zhuang Autonomous Region, and Hainan Province.17 These areas are geographically distant, and farming management practices and environmental conditions differ from location to location. This study investigated morphological differences among seven different geographic populations of golden pompano, and the results showed that the degree of difference varied among different populations. Although the morphological differences between some populations were significant, there was still some variation among them, and it was impossible to use this basic morphological analysis to identify all populations. This indicates that the differences resulting from this morphological variation are unstable. To some extent, stable morphological differences can reveal population growth, mortality, and reproductive rates.18
Single-factor variance analysis (Table 5) showed that different parts of the body that reflected the differences among different populations were different. Although there were differences in the six indicators between the HX and LL populations, JG and LL populations, LL and DL populations, and JG and NH populations, there was no significant difference in body depth and length of caudal peduncle between the HX and LL populations, no significant difference in body depth and head length between JG and LL populations, no significant difference in head length and snout length between LL and DL populations, and no significant difference in head length and eye diameter between JG and NH populations. Discriminant analysis of eight proportional traits showed the overall accuracy of the samples classified into the CH and DL populations was the highest. In contrast, the classification rate of the NH population was the lowest, which may be related to its slight morphological variation. The comprehensive discrimination rate based on the discriminant formula established from the eight proportional traits was 71.4%, indicating that using the formula to differentiate group characteristics is reliable for many well-preserved and differently-sized golden pompano samples.
In early research, the differences in morphological characteristics among populations were often attributed to genetic factors,19 but genetics only accounted for the majority. Body shape is a product of individual development,6 and studies have shown that the morphological characteristics of fish are more susceptible to the influence of external environmental factors among vertebrates. Therefore, environmental factors’ influence on fish growth and development cannot be ignored, and variation in morphology has environmental and genetic influences.20–22 In this study, the HX and DL populations showed the greatest morphological differences, with all eight morphological indicators showing significant differences. Their living environments were different. Hainan Island is located at the southernmost tip of China, with slightly higher water temperatures than the Zhanjiang area. The waters near Hainan Island are more open, with better biological abundance, water conditions, and dissolved oxygen, so this living environment may exert different selection pressures on these two populations. Franssen et al. reported that the similarity in external morphological characteristics might be due to insufficient selective pressure from fish living environments,23 making the effect of genetics more pronounced. Principal component analysis (PCA), known as the Karhunan-Loeve transform, is an effective data analysis method. This analytical approach can use a few features to describe the training samples, reducing the dimensionality of multi-dimensional data.24,25 Therefore, PCA exhibits the characteristics of dimensionality reduction, correlation removal, and energy concentration. The scatter plot of the principal components in this study showed overlap between the different populations, possibly due to weak environmental pressure and genetic effects. The influence of genetic differentiation on phenotypic morphological differences cannot be ignored; however, the strength of the environmental impact and genetic differentiation requires further studies. Zhang et al. studied the genetic differentiation of the Chinese Gizzard Shad Clupanodon thrissa using morphological and molecular markers. Results showed no significant morphological differences between them, but there was significant genetic differentiation.26 At the same time, some scholars have proposed that the morphological differences between species, especially subspecies, are consistent with molecular differences. 27,28
From the hatching of fertilized eggs to the death of fish, environmental pressure plays a constant role in life history. Meng and Austin et al. suggested that morphological differences among different populations might also be due to environmental differences during spawning and juvenile fish development.29 The larval and juvenile stages are critical periods in fish development; thus, environmental pressure may be greater. Huang et al. found a significant correlation between body length, depth, and weight during the early development of the golden pompano.30 Therefore, this study suggests that when identifying different geographical populations of the golden pompano using traditional morphological data, it is essential to consider more features related to body length and depth. Targeted management strategies based on differences in body length can aid the breeding of golden pompano seedlings.
It is challenging to achieve precise results when using the combined approach of morphology and multivariate statistics to identify different populations in this study. One reason for this is that the sample sizes of different populations vary. During data processing, morphological feature data are usually normalized by their body length or total length ratio as the trait value for multivariate statistical analysis. This method was applied to the raw data used in this study. However, even for the same fish population, the range of phenotypic morphological ratios can differ at different developmental stages. Therefore, this approach only partially reduced population differences and did not eliminate the influence of different fish sizes on the research results.
Analyzing the morphological differences among different populations helps understand the population differences and structure of the golden pompano. It can aid in developing management and breeding strategies targeted at different ecological populations. This study demonstrates that, although the morphological approach can identify some golden pompano populations, a more accurate distinction of different populations requires a combination of molecular biology methods for a more scientifically precise judgment.
Conclusions
This study combined external morphological features and diversified statistical methods to analyze different geographical populations of golden pompano and found differences in their morphological characteristics. The first three principal components could better explain the original eight proportional traits. NH, CH, and LL populations had the most similar forms, whereas the HX and DL populations had the greatest differences in form. There were specific differences in certain traits between other populations, and the degree of differentiation of the populations could not be identified by basic morphological analysis. The discriminant analysis showed an overall high accuracy, with the DL population having the highest discrimination accuracy. The study of fish morphology for different geographical populations provides new perspectives for understanding the spatial structure of populations and developing targeted farming management strategies. In the future, it will be necessary to jointly explain the mechanisms of morphological differentiation of different geographical populations from environmental effects and genetics perspectives.
Acknowledgments
This research was financially supported by the Key areas of Guangdong province research and development projects (2021B0202020002), China Agriculture Research System (CARS-47), the National Natural Science Foundation of China (U20A2064), National Marine Genetic Resource Center.