




Introduction
Reactive oxygen species (ROS, such as O2-, OH-, and H2O2) actively guard bacteria in the host or function as signaling chemicals in various inflammatory illnesses or innate immune responses.1 The complex antioxidant defense system, one of the primary adaptive strategies against pathogen invasion in fishes, can eliminate the overproduction of ROS. Microbial elicitors may promote oxidative bursts and accelerate the production of reactive oxygen species (ROS), which combat and kill invading microorganisms.2 Nonetheless, the mass accumulation of ROS in vivo would reduce antioxidant defenses or cause oxidative damage to biological macromolecules, causing various disorders.3 Organisms have evolved complex and integrated antioxidant defense systems that include superoxide dismutases (SODs), catalase (CAT), and different peroxidases to prevent the harmful effects of reactive oxygen species (ROS).4
SODs are essential for managing oxidative damage and the first line of defense against ROS.5 CAT converts superoxide radicals into less reactive H2O2, which is then catalyzed into H2O by SODs.6 SODs typically consist of four varieties based on subcellular localization and bound iron: FeSOD, MnSOD, Cu/Zn-SOD, and NiSOD.7 MnSOD is particularly important for maintaining cellular homeostasis and removing excessive ROS from the host. CAT is essential for detoxifying H2O2 into the non-toxic end products H2O and O2.8 SOD and CAT can eliminate excessive ROS to maintain oxidation-antioxidation equilibrium and protect animals from oxidative injury.9 In addition, MnSOD and CAT can regulate mRNA transcription, which helps to understand the mRNA expressions of MnSOD and CAT to combat infections and provides fresh information regarding fish acclimation.
Since its introduction to Northern China in 1992, turbot (Scophthalmus maximus), one of the most frequent mariculture species with rapid growth and high resistance to low water temperatures, has been vital to fisheries’ economic and aquaculture species. However, infections caused by bacteria, viruses, and parasites (such as Vibrio anguillarum, nodavirus, and Philasterides dicentrarchi) in booming turbot cultures have caused significant economic losses for the turbot business.10 According to reports, V. anguillarum is mainly responsible for severe infections in cultured turbot. Thus, additional research into the innate immune capabilities and the immune defense mechanism against V. anguillarum will contribute to disease control and turbot culture’s chronic survival. This study aims to describe the nucleotide sequence of MnSOD from the liver of S. maximus in comparison to other known MnSODs, detect the expressions of MnSOD in various tissues, and quantify the expressions of MnSOD and CAT in response to V. anguillarum. Our findings would contribute to understanding the characteristics of MnSOD and CAT in fish.
Materials and methods
Animal materials
Juvenile turbot weighing approximately 45.05±2.16 g were acquired from a farm in Changli, China. Scophthalmus maximus was first acclimated to the experimental conditions by spending one week in aerated, clean saltwater (salinity 19‰, 16±1 °C). Each group consisted of ten turbots and three fish tanks served as replicas. During the acclimation period, the turbots were fed twice daily with commercial pellets (vendor) with continuous oxygenation, and one-fourth of each tank’s saltwater was replaced daily.
Total RNA isolation from turbot liver and cDNA preparation
Total liver RNA was isolated directly from healthy turbot using an RNAiso Plus system (TaKaRa, Japan). RNA’s integrity and quantity/quality were then determined using agarose electrophoresis and an ND-1000UV (NanoDrop, USA) spectrophotometer, respectively. All extracted samples had an OD260/OD280 ratio between 1.8 and 2.0. Using M-MLV reverse transcriptase, first-strand cDNA was generated (TaKaRa). Using a SMART™ cDNA Kit, the 5′ and 3′ rapid amplification of cDNA end (RACE) cDNA templates were obtained (Clontech, USA). The manufacturer’s instructions carried out all procedures.
Cloning of S. maximus MnSOD
S. maximus MnSOD (SmMnSOD) full-length cDNA was produced using reverse-transcription polymerase chain reaction (RT-PCR) and RACE. The middle fragment of the SmMnSOD cDNA sequence was identified using degeneration primers F1 and R3 (Table 1) for the conserved regions and by aligning with the target genes of other fish species, including Rachycentron canadum, Sparus aurata, Paralichthys olivaceus, Oplegnathus fasciatus, Anoplopoma fimbria, and Larimichthys crocea. The PCR conditions were as follows: denaturation at 95°C for 5 minutes, followed by 33 cycles of denaturation at 95°C for 40 seconds, annealing at 56°C for 40 seconds, elongation at 72°C for 1 minute and 10 seconds, elongation at 72°C for 10 minutes, and storage at 4°C. Finally, a 463-bp fragment of MnSOD cDNA was amplified.
Using SMART™ RACE cDNA Kit, the 5′ and 3′ ends of the preceding SmMnSOD fragment sequence were obtained. The cDNA template derived from liver RNA, GSP-R1, and the anchor universal primer (UPM, Table 1) were utilized in the PCR of 5′ end RACE. The GSP-F1 and the anchor UPM (Table 1) were selected for the 3’ end of the RACE. In the SMART cDNA Kit manual, the PCR settings were specified.
The target band was excised and purified using a gel isolation kit (TaKaRa) before being subcloned into the pMD18-T cloning vector according to the manufacturer’s instructions (TaKaRa) and transformed into competent Escherichia coli DH5. After PCR validation, Sangon Biotechnology Co., Ltd. sequenced the positive clones containing the DNA insert (Beijing, China).
Sequence investigation
The nucleotide and estimated amino acid sequences of SmMnSOD cDNA were compared using BLAST (http://www.blast.ncbi.nlm.nih.gov/Blast), SignalP 4.1 (http://www.cbs.dtu.dk/services/ SignalP/), and ExPASy tools (http://us.expasy.org/tools/). DNAMAN was used for multiple alignment of the turbot MnSOD gene (Lynnon Biosoft, Canada). MEGA 11 software was used to create a phylogenetic tree of MnSOD, and statistical support for reliability was assessed using 1000 bootstrap replications.11
Differential expressions of SmMnSOD detected by RT-qPCR
Primer Premier 5.0 was used to construct the qRT-PCR primers QF, QR, and -actin-F/R (Table 1) based on the SmMnSOD gene sequence and the internal housekeeping control gene β-actin (GenBank number: AY008305). Total RNA samples (1g) from healthy S. maximus tissues (gill, head, kidney, liver, stomach, muscle, heart, and intestine) were reverse-transcribed into cDNA using gDNA Eraser (TaKaRa) according to the manufacturer’s protocol, and the expression levels of MnSOD mRNA transcript in each tested tissue were assessed using quantitative real time PCR (qRT-PCR).
Expression patterns of SmMnSOD and CAT mRNA after V. anguillarum injection
The turbots were divided into two groups: test and control. Each turbot was injected intramuscularly with 100 μL of a live bacterial suspension (2×107 CFU/mL) of V. anguillarum strains in the test group, which had been obtained from diseased S. maximus in Longxiang aquaculture farm (Laoting, China). Instead, sterile PBS was used in the control group. Following this treatment, the lives and kidneys of three turbots from each group were randomly taken at 3, 6, 12, 24, 48, and 72 h, and the samples were snap-frozen in liquid nitrogen for total RNA extraction and first-strand cDNA synthesis, as described above. Each time point was tested three times.
SmMnSOD and CAT (GenBank number: MG253621) post-injection expressions were detected using a CFX96 RT-PCR system (Bio-Rad, America) and SYBR green I. For SmMnSOD, a 162-bp PCR product was amplified using the gene-specific primers QF and QR. Still, for CAT, a 187-bp PCR product was amplified first with the specific primers QF and QR, and then the endogenous reference gene β-actin was amplified as a 203-bp product using the primers β-actin-F and β-actin-R (Table 1).
The cDNA was synthesized with a PrimeScript™ RT reagent kit with gDNA Eraser (TaKaRa, Japan) using 1 μg of total RNA. The constructed cDNA was diluted 10-fold in DEPC water and stored at −70 °C for qRT-PCR. qRT-PCR was performed in triplicate in a total volume of 10 μL, containing 1× SYBR Premix Ex Taq™ II (TaKaRa), 0.4 μM of each primer, and 1 µL of an RT solution. The settings for PCR amplification were provided in the manufacturer’s instructions. Following each PCR thermal profile, the threshold cycle (CT) was calculated using CFX96™ real-time PCR software 1.0 and applied to calculate the ΔCT of each sample.12 2-ΔΔCT was used to determine the relative expression of SmMnSOD and CAT.13
The mean, standard deviation (n=3) is presented for all experimental results. The qRT-PCR results were analyzed using a one-way analysis of variance (ANOVA) in SPSS 17.0 software. P < 0.05. was used as the significance level.
Results
Characteristics and bioinformatic analysis of SmMnSOD sequence
RACE-derived full-length SmMnSOD cDNA (Figure 1) was uploaded to the Genbank database (Accession Number: MG253620). As predicted by SignalP 4.1, the 1267-bp SmMnSOD cDNA has a 684-bp open reading frame (ORF) that encodes a 228-amino-acid protein with a 28-amino-acid signal peptide (Figure 1). The computed molecular mass of the protein was 23.87 kDa, and the theoretical isoelectric point was 6.75. The SmMnSOD amino acid sequence contained putative N-linked glycosylation sites of NVT, NHT, and NLS, as well as highly conserved signatures (DVWEHAYY).

Homology analysis of SmMnSOD
According to sequence analysis on the BLASTP, the deduced sequence of SmMnSOD had the same consensus pattern as the MnSODs of vertebrates and had an overall identity of 85.3 % to 92.9 % with eight different known MnSOD members (Figure 2). The highest identity was observed with homologs from Oplegnathus fasciatus and Rachycentron canadum, with 92.9 % identity.

Phylogenetic analysis
The identification and phylogeny of SmMnSOD were confirmed further using a phylogenetic tree constructed on MEGA 11 using the sequences of the selected MnSODs (Figure 3). The MnSOD gene family is thought to be divided into two primary phylogenic clusters. The SmMnSOD was similar to some fish species but not to Homo sapiens, Gallus gallus, Melopsittacus undulatus, or Mus musculus MnSODs (Figure 3).

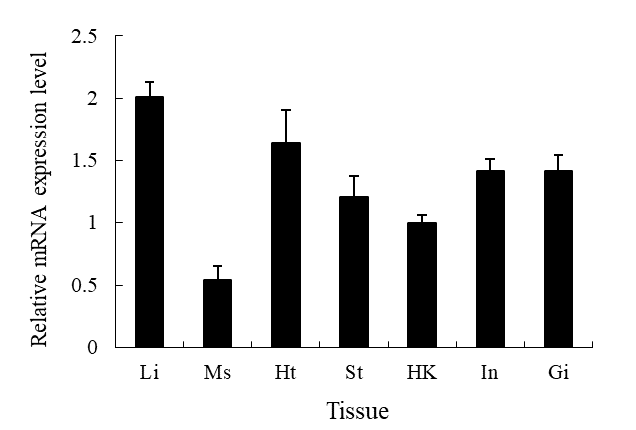
Basal tissue expression of SmMnSOD mRNA
SmMnSOD gene expression patterns were studied in seven different tissue types of healthy turbot, including the gills, stomach, liver, head, kidney, intestine, heart, and muscle (Figure 4). SmMnSOD and β-actin dissociation curves only showed one peak, indicating that the amplifications were unique. The results showed SmMnSOD expression was highest in the liver, lowest in the muscle, and moderate in the other tissues (Figure 4).
__ms_(muscle)__ht_(heart)__st_(stomach).png)
Expression profiles of SmMnSOD after V. anguillarum challenge
The transcript levels of SmMnSOD in the liver and head kidney following V. anguillarum infection were measured using qRT-PCR with the β-actin gene as the endogenous reference. Compared to the control group, SmMnSOD transcript levels increased considerably in the liver from 12 to 72 hours (P<0.05), maximized at 24 hours with an 8.9-fold increase for the challenge group (Figure 5a).
_and_the_head_kidney_(b)_of_*s._maximus*_afte.png)
SmMnSOD mRNA expression in the test group’s head kidney declined quickly after the challenge, and was significantly lower than in the control group during the first 12 hours (P<0.05; Figure 5b). Following that, SmMnSOD mRNA expression increased progressively in the challenge group, maximized at 48 h with a 2.3-fold increase relative to the control group (P<0.05), and subsequently decreasing to 1.29-fold at 72 h (P>0.05) (Figure 5b).
Expression profiles of CAT after V. anguillarum challenge
Figure 6 shows the CAT expression profiles following pathogenic infection. In the challenge group, CAT expression in the liver of S. maximus increased steadily from 3 to 48 h, maximized at 48 h with a 4.17-fold increase compared to the control group (P<0.05). The CAT transcript level in the liver then reduced after 72 hours but remained significantly more significant than in the control group (P<0.05; Figure 6a).
_and_the_head_kidney_(b)_of_*s._maximus*_after_th.png)
Within the first 6 hours, the CAT transcript level in the head kidney of S. maximus was significantly lower than the control group (P<0.05). The CAT expression level then steadily increased in the challenge group, although it increased significantly from 24 to 72 h, maximized at 48 h by 1.59-fold compared to the control group (P<0.05; Figure 6b).
Discussion
SODs are important metalloenzymes in antioxidant defense pathways that reduce ROS buildup and free-radical-caused cell damage in vivo.14 SODs are the first line of defense against oxidative stress caused by massive ROS levels in the body. In this article, for the first time, a novel 1267-bp full-length MnSOD gene was isolated and described from S. maximus, and it showed high similarity (85.3 % - 92.9 %) with MnSODs from other animals. Multiple sequence alignment revealed that the SmMnSOD hallmark motifs (DVWEHAYY) were substantially conserved in species15 (Figure 2). Phylogenetic research revealed that SmMnSOD belonged to the fish SOD family and clustered with fish homologs. The presence of MnSODs in animals shows that MnSODs have a common ancestor and are conserved during evolution. The SmMnSOD belongs to the MnSOD family based on its close relationship with other fishes.
SmMnSOD was shown to be expressed in all turbot tissues tested using qRT-PCR. The SmMnSOD transcript expression was highest in the liver, which is consistent with previous findings in Megalobrama amblycephala,16 Hypophthalmichthys molitrix,17 and Hemibarbus mylodon.17,18 However, this contradicts another finding showing MnSOD expression is highest in the heart of Sebastes schligellii.19 Furthermore, the lowest expression level in muscle in our investigation contradicts Hemibarbus mylodon.18 MnSOD expression patterns in fish may be attributed to the tissue dependence on mitochondrial and oxidative levels, as MnSOD primarily deletes ROS generated during mitochondrial respiration.18 According to findings, antioxidant enzymes are expressed at the mRNA level in the liver of many fish species.20 SmMnSOD transcript levels are highest in the liver, most likely due to oxidative reactions and highly metabolic antioxidant defenses that always occur in the liver.21 These findings indicate that MnSOD in the liver plays a vital role in antioxidant defenses.
Virulent pathogens have a significant impact on aquaculture development. The experimental challenge of pathogen bacteria significantly affects MnSOD expression in Takifugu obscurus, Pseudosciaena crocea, Hemibarbus mylodon, Procambarus clarkia, Crassostrea hongkongensis, and Meretrix meretrix. V. anguillarum, a significant pathogen bacterium, is said to cause severe infections and widespread mortality in farmed marine fishes.22 There has yet to be any research on the effect of V. anguillarum on MnSOD expression in one of its most heavily attacked species, S. maximus.23
Since the liver and kidney are the primary sites of oxidative reactions and antioxidant defenses,24 determining the MnSOD gene expression levels in these two tissues following V. anguillarum infection provide more insight into its immunological function. According to our findings, Live V. anguillarum strongly activated MnSOD in the liver and head kidney of S. maximus. SmMnSOD expression increased considerably after 48 h in the test group against the control group, which might be attributable to the ROS stress generated by V. anguillarum infection, indicating that SmMnSOD is engaged in antioxidant defense and can be induced when infected with V. anguillarum. SmMnSOD expression in the liver and head kidney gradually decreased at 24 and 48 hours, respectively. MnSOD transcription was inhibited in the challenged group, most likely because the continuous reproduction of viable V. anguillarum in hosts created severe cytotoxic effects and severely impaired normal cell function, resulting in either apoptosis or cell-function loss.25,26
These findings suggest that MnSOD may play a role in the immunological response against V. anguillarum infection. We also found that MnSOD expression patterns differed between the liver and the head kidney, most likely due to the antioxidant defense system’s various cell and tissue activities.27 Another MnSOD expression investigation from Carassius auratus yielded similar results.28 The transcript profile analysis in our study shows that pathogenic bacteria can induce MnSOD, implying that MnSOD plays an essential role in S. maximus’ antioxidant defense response. Because MnSOD is a component of the antioxidant system, monitoring its expression alone cannot provide a complete picture of the oxidative stress caused by pathogenic bacteria. Thus, additional CAT research is required for ecotoxicity monitoring. The explanation is that SOD and CAT mitigate the adverse effects of ROS by converting them into O2 and H2O, thereby protecting organisms from lipid peroxidation and balancing cellular redox reactions.17,29
Past studies have shown that pathogenic bacteria such as Vibrio alginolyticus, Streptococcus iniae, and V. alginolyticus can produce CAT quickly. In our study, V. anguillarum dramatically increased CAT mRNA transcription levels in the liver and head kidney at 6 and 24 h, respectively, peaked at 48 h, and then progressively reduced, indicating that CAT was inducible and likely participates in antipathogenic immunological reactions in S. maximus. Unfortunately, it has not yet been determined why the induction of MnSOD and CAT by V. anguillarum challenge is more significant in the liver than in the head kidney. One probable explanation is that the liver is the principal organ involved in oxidative reactions and antioxidative defenses.18 Nonetheless, more research is needed to determine the precise mechanism of bacterial infection on antioxidant genes in S. maximus.
In summary, MnSOD was identified in S. maximus for the first time, and it was found to be constitutively expressed in the liver (highest), gills, stomach, head-kidney, muscle, intestine, and heart. MnSOD and CAT expressions in the liver and head kidney following V. anguillarum challenge revealed that MnSOD and CAT were inducible and may have engaged in the innate immune response of fishes against pathogenic infection. This study provides some fresh and significant evidence for further research into the regulatory mechanism of the antioxidant defense system against infections in flounder fishes.
Acknowledgments
Financial support from the National Natural Science Foundation of China (31502187), the Major Science and Technology Projects of Hebei Province (20286701Z), the Natural Science Foundation of Hebei Province (C2018407049), Science and Research Funds of Hebei Normal University of Science and Technology (2020YB008), the Science and Technology Projects of Hebei Province (20567621H) are gratefully acknowledged. The authors thank EditSprings (https://www.editsprings.cn) for the expert linguistic services provided.
Declaration of Competing Interest
The authors have stated that no competing interests exist.
Authors’ Contribution - CRediT
Conceptualization: Hai Ren (Equal), Shaoyu Yang (Equal). Methodology: Hai Ren (Lead), Jing Mei (Supporting). Formal Analysis: Hai Ren (Lead). Investigation: Hai Ren (Lead). Writing – original draft: Hai Ren (Lead). Funding acquisition: Hai Ren (Lead). Writing – review & editing: Qinglin Wang (Supporting), Xiaomin Jin (Supporting), Guisheng Gao (Equal). Resources: Guoshan Qi (Equal), zenping He (Equal). Supervision: Shaoyu Yang (Lead).