Introduction
The green crucian carp (Carassius auratus indigentiaus subsp. nov.) is a new subspecies of crucian discovered by our research team during a resource survey in Dongting Lake, China. It is a unique species that remains in the marginal waters of this lake.1 Preliminarily studies of its morphological features, karyotype, nutrition, growth, and genetic diversity2 indicate that this crucian carp subspecies has digenetic propagation and a stable chromosome number. It also has molecular genetic differences compared to other crucian carp species.3–5 Green crucian carp is a new member of the carp family and provides good experimental material for discussing the genetic relationships among different crucian carp species. It has great theoretical significance for disclosing the evolutionary mechanisms of this crucian carp subspecies in nature.
When cultivated in the same pool, the growth rate of green crucian carp is at least 10% higher than those of its conspecifics C. a. pengzesis and C. a. cuvieri. The highest weight of one-year-old individuals is 400 g, and the average weight of two-year-old is > 430 g. The maximum adult weight can reach 1221 g. The capture rate can be as high as 95%. In green crucian carp, the flavor amino acid accounts for 50% of total amino acids, and its water content is about 1%. The new species has the potential for aquaculture, having a good taste, rich nutrient profile, and stable inheritable traits. However, the early embryonic development of green crucian carp has not yet been studied. Through systematic observation, photographic records and analysis, this study revealed green crucian carp’s early embryonic developmental characteristics, laying a foundation for its large-scale aquaculture.
Embryonic development is the most critical stage of fish development and is affected by many external factors such as temperature, pH, salinity, and dissolved oxygen concentration.6 Temperature is one of the most important factors because animals must absorb a certain amount of heat from the environment to complete growth and development. The total heat required for each stage of development is constant; that is, the total accumulated temperature. Therefore, in the appropriate temperature range, the time required for each stage of embryonic development in fish will decrease with temperature increases. In addition, the early development of fish is also very sensitive to temperature: too high or too low can lead to abnormal embryo development or death.7,8 Synodontis embryo development speeds up with temperature within a suitable range; for example, the total developmental time is shortened by 13.1–25.3% with a 2 °C increase in water temperature.9 Above the optimum temperature, the embryo hatching rate decreases while the malformation and mortality rates increase.10
There are several classifications of embryonic development in fish. Some scholars have proposed that development from a fertilized egg to a larva can divided into three major stages: cleavage, blastogastrula formation, and tissue and organ formation. Morphogenesis and organogenesis in grass carp embryonic development have been divided into six stages. This view is widely adopted and adapted to different fish’s development. In cyprinid fishes, the time series of embryonic development has been studied thoroughly, including in the hybrid of Caratus auratus gibelio × Cyprinus carpio,11 Carassius auratus pengzesis,12 Cymnocypris chilianensis,13 Cyprinus carpio haematopterus,14 Gymnodiptychus pachycheilus,15 Schizothorax chongi,16 Salmo trutta macrostigma and Salmo trutta labrax,17 Paracanthobrama guichenoti,18 triploid crucian carp,19 Carassius auratus gibelio,20 Lyprinus carpio Linné,21 the hybrid of Ctenopharyngodon idellus♀ × Elopichthys bam busa♂,22 zebrafish,23 Acrossocheilus fasciatus24 and Tinca tinca Linnaeus.25 Based on the above studies, it can be concluded that the embryonic development of cyprinid fishes can generally be divided into a blastoderm formation stage, mitotic stage, blastula stage, gastrula stage, neurula stage, organogenetic stage, and hatching stage. There are distinct phenomena in each stage. For example, the protoplasm concentrates toward the yolk end, uplifting the blastoderm, forming gradually during the blastoderm formation stage. However, different cyprinid fishes display different developmental phenomena, such as the timing and duration of different stages and the developmental order of different organs. In this study, the early embryonic development of green crucian carp was observed systematically to provide a reference for its breeding and cultivation.
Materials and Methods
Selection of parent fish
The parent fish used in the experiment were 1–2-year-old wild green crucian carp captured at Dongting Lake. Healthy individuals without malformation, smooth bodies, scales, and complete fins were selected. Acceptable female parent fish were those with an expanding, soft, and elastic belly, while male parent fish were those that released seminal fluid upon squeezing the abdomen. The experimental water temperature was 24–26 °C.
Artificial insemination and embryo observation
The hormone oxytocin was injected into experimental parent fish to induce spawning. Oxytocin uses the DOM and Luteinizing hormone-releasing hormone analog (LRH-A2). The injection dosage for female fish was DOM 1 mg/kg + LRH-A2 5 µg/kg, while that for males was 50% less. Dry fertilization by egg squeezing was performed at 12 hr at a water temperature of 24 ± 1 °C, salinity of 35 ± 1, and pH of 7.4 ± 0.5. Since the eggs of green crucian carp are viscous, fertilized eggs used for embryonic development were arranged uniformly on a 16 cm-diameter petri dish for the convenience of sampling and observation. The water was changed every 3 hr to ensure that the dissolved oxygen content was acceptable for the normal development of embryos. During embryonic development observations, 30 fertilized eggs were collected and observed under a microscope after 3–5 min. The petri dishes and incubator were kept clean throughout the experiment, and dead eggs were eliminated promptly to avoid affecting the growth and development of other fertilized eggs. The effective accumulative temperature was calculated as °C × hr, where °C is the average nychthemeron water temperature and hr is the number of development hours in that stage. The observation was performed under a Huaca stereoscope, and pictures were taken using LAS V4.5 (Leica Application suite). Embryonic development stages were defined according to > 50% of the individuals presenting the characteristics of that stage.
After hatching, six larvae were taken at 10:00 each day to observe their growth and development characteristics with an anatomical microscope. The morphological changes and organ development of larvae and juveniles were observed, and their full length, total length, body height, head length, eye diameter, and preanal distance were measured. Data were analyzed using Excel 2020 software, and regression equations describing the growth characteristics of juvenile fish at different stages were calculated.
Results
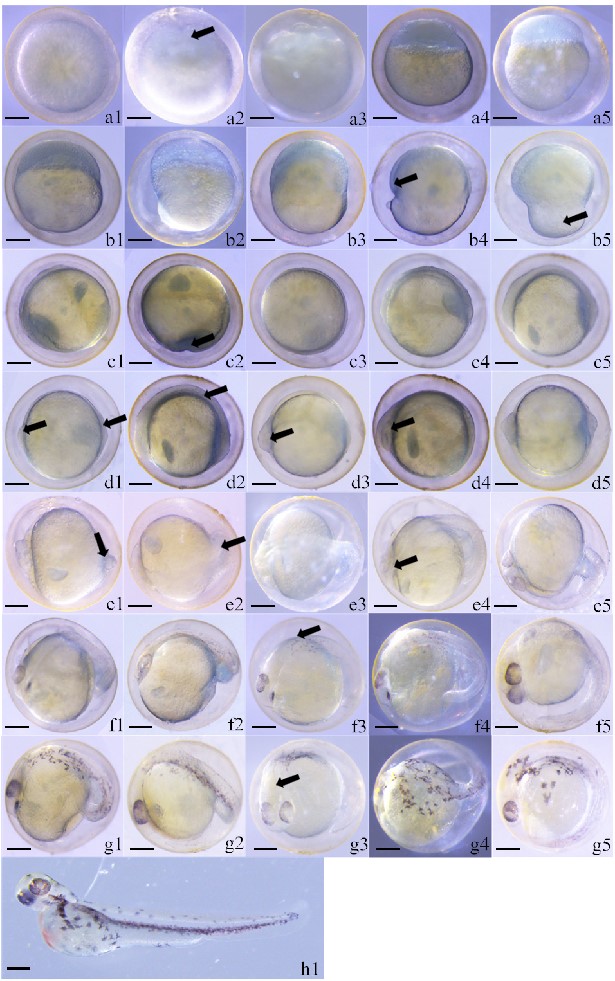
According to the observations, the early embryonic development of green crucian carp lasted 56.5 hr. It can be divided into the following stages: blastoderm formation stage, mitotic stage, blastula stage, gastrula stage, neurula stage, blastopore closure stage, organogenetic stage, and hatching stage. The first seven stages lasted for 55 min, 165 min, 240 min, 215 min, 25 min, 50 min and 2640 min, respectively. The effective accumulative temperature from fertilization to membrane formation was 1361.7 °C x hr (Table 1).
Blastoderm formation stage
Fertilized eggs of green crucian carp are round, light yellow, and viscous. They swell due to water absorption. The egg membrane is separated from the yolk, and the egg membrane is strong and difficult to break. After water absorption, the diameter of fertilized eggs was 1.47 ± 0.04 mm (Figure 1 a1). The blastoderm began to form 35 min after fertilization. At this moment, the protoplasm began to concentrate toward the animal pole of the egg.

Mitotic stage
At 55 min, fertilized eggs began to form a cleavage furrow and entered the 2-cell phase (Figure 1 a2), then the 4-cell, 8-cell (Figure 1 a3), 16-cell, and 32-cell phases. They continued to split 170 min after fertilization, then entered the multi-cell phase (Figure 1 a4). The number of cells increased continuously while the cell volume decreased continuously. Moreover, the cell size varied, increasing the number of cell layers. Mild fluctuations in the yolk, from depressions to bulges, could be observed in this stage. As fluctuations of the egg yolk continued, it became irregular but maintained its orientation.
Blastula stage
At 220 min, the blastoderm continued to increase with ongoing merogenesis. A semi-circular uplift with smooth edges, the blastaea, formed on the yolk. The uplift height accounted for 1/3 of the yolk (Figure 1 a5). Subsequently, the blastomere wrapped downward along the yolk, and the blastaea height declined accordingly. At 230 min, the blastomere was wrapped downward to 1/5 of the yolk sac, and the middle blastula phase began (Figure 1 b1). At 245 min, the blastomere was wrapped downward over 1/3 of the yolk sac, and the late blastula phase began (Figure 1 b2).
Gastrula stage
At 460 min, blastomere wrapped downward over half of the yolk in a C-shape. The early gastrula phase then began (Figure 1 b3). At 470 min, cells continued to wrap downward, and a semi-annular region appeared on the yolk. The embryo was flattened, and the part wrapping the yolk had a consistent height. This structure is called the germ ring. As the germ ring formed, embryonic development entered the middle gastrula phase (Figure 1 b4). At 610 min, the germ ring cells were wrapped downward over about 3/4 of the yolk. The part not enclosed by the germ ring is called the yolk plug. The blastomere then entered the late gastrula phase (Figure 1 b5).
Neurula stage
At 675 min, the blastomere was wrapped downward over 9/10 of the yolk. This is the neurula stage. The yolk did not fluctuate, recovered to a static state, and was round (Figure 1 c1). At 690 min, almost the embryo was wrapped, and only a small hole was left, called the blastopore (Figure 1 c2).
Blastopore closure stage
At 700 min, the blastopore closed, and the germ ring was very close to the yolk. It formed a concentric-circular structure with the yolk as the inner circle and the germ ring as the outer ring (Figure 1 c3).
Organogenetic stage
At 750 min, a part of the germ ring protruded to form an embryonic shield. The idiosome was a relatively thick, solid, and non-transparent arc close to the yolk. The idiosome rounded the yolk for about 2/3 of a circle and laid on the side (Figure 1 c4). At 755 min, the idiosome was obvious, and the embryonic shield thickened significantly, especially at the two ends (Figure 1 c5). At this moment, the head and tail could be distinguished clearly, the head being raised high and the tail relatively flat (Figure 1 d1). At 815 min, the idiosome began to develop a short, columnar sarcomere near the middle part, which developed five pairs (Figure 1 d2). At 900 min, the head was raised, and eye primordia developed at the two sides of the brain vesicles. However, the eye crystal had not yet developed. The eye primordium seemed oval, and a seam could be seen indistinctly in the middle. The tail became even flatter (Figure 1 d3). At 910 min, the eye primordia and the middle seam were more obvious. The sarcomere developed at the tail, and the middle idiosome was relatively flat. Most embryos still lay on the side, and the idiosome extended gradually (Figure 1 d4). At 1000 min, it could be seen from the segmental venter that the yolk was not oval anymore. The yolk at the head and tail narrowed, and the tail was raised (Figure 1 d5). At 1200 min, the yolk sac was approximately inverted and pear-shaped, and the cardiocoelom was preliminarily formed. Spherical bubbles were seen at the tail (Figure 1 e1). At 1210 min, the tail separated from the yolk sac and curled up. The tail bud began to form, which looked like an inverted cone shape (Figure 1 e2). At 1300 min, the tail swung slightly (Figure 1 e3). At 1305 min, slightly grey eye pigments began to appear.
Meanwhile, eye crystals began to appear, which were round. Under this circumstance, the tail fin primordium could be seen indistinctly and was almost transparent. The tail could swing horizontally to a small extent (Figure 1 e4). At 1315 min, shaped eyecups were visible (Figure 1 e5). At 1394 min, the black eye pigments increased abundance and gradually changed from light grey to black, accompanied by an obvious eye crystal. The eye sac became increasingly round (Figure 1 f1). At 1415 min, the eye crystal became more obvious (Figure 1 f2). At 1490 min, black pigments appeared on the back (Figure 1 f3). At 1630 min, body pigments dispersed to the yolk, and the idiosome twisted frequently; this rapid twisting could change the positions and directions of the idiosome in the egg membrane (Figure 1 f4). At 1650 min, the idiosome could rotate in the egg membrane, and the tail was long enough to reach the head (Figure 1 f5). Pigments spread out at 1765 min, and the tail separated from the yolk sac completely. Tail pigments were formed, and there was a deep depression at the lower part of the yolk sac (Figure 1 g1). At 1765 min, blood flowed in the cardiocoelom (Figure 1 g2). At 1860 min, the ventricle could be seen very clearly (Figure 1 g3). At 1985 min, pigments spread to the yolk sac (Figure 1 g4). At 2700 min, violent heartbeats in a bag-shaped could be seen in the cardiocoelom with light-red blood (Figure 1 g5).
Hatching stage
At 3390 min, fish hatched out of the membrane tail-first. The fish lay on their side after hatching. The whole fish body was covered with pigment except for the yolk and head, especially surrounding the spine. Fish developed a pectoral fin and tail fin at hatching. Moreover, the eyes had colorful pigments, but the eye crystals were colorless. The eyes occupied a high proportion of the whole head area and were approximately spherical. The yolk sac had not disappeared completely, and the abdominal yolk sac swelled. The tail yolk sac was relatively slim, and the vertebra was obvious, accompanied by obvious heartbeats. There were obvious blood circulation lines on the belly surface of the yolk sac. Left and right blood vessels were seen clearly on the yolk. Blood concentrated towards the heart, and a blood vessel was along the spine. Blood was bright red, and the cloacal opening had formed. A simple digestive tract and a pair of bright otoliths were behind the eyes, but the mouth was not yet opened (Figure 1 h1).
Growth characteristics
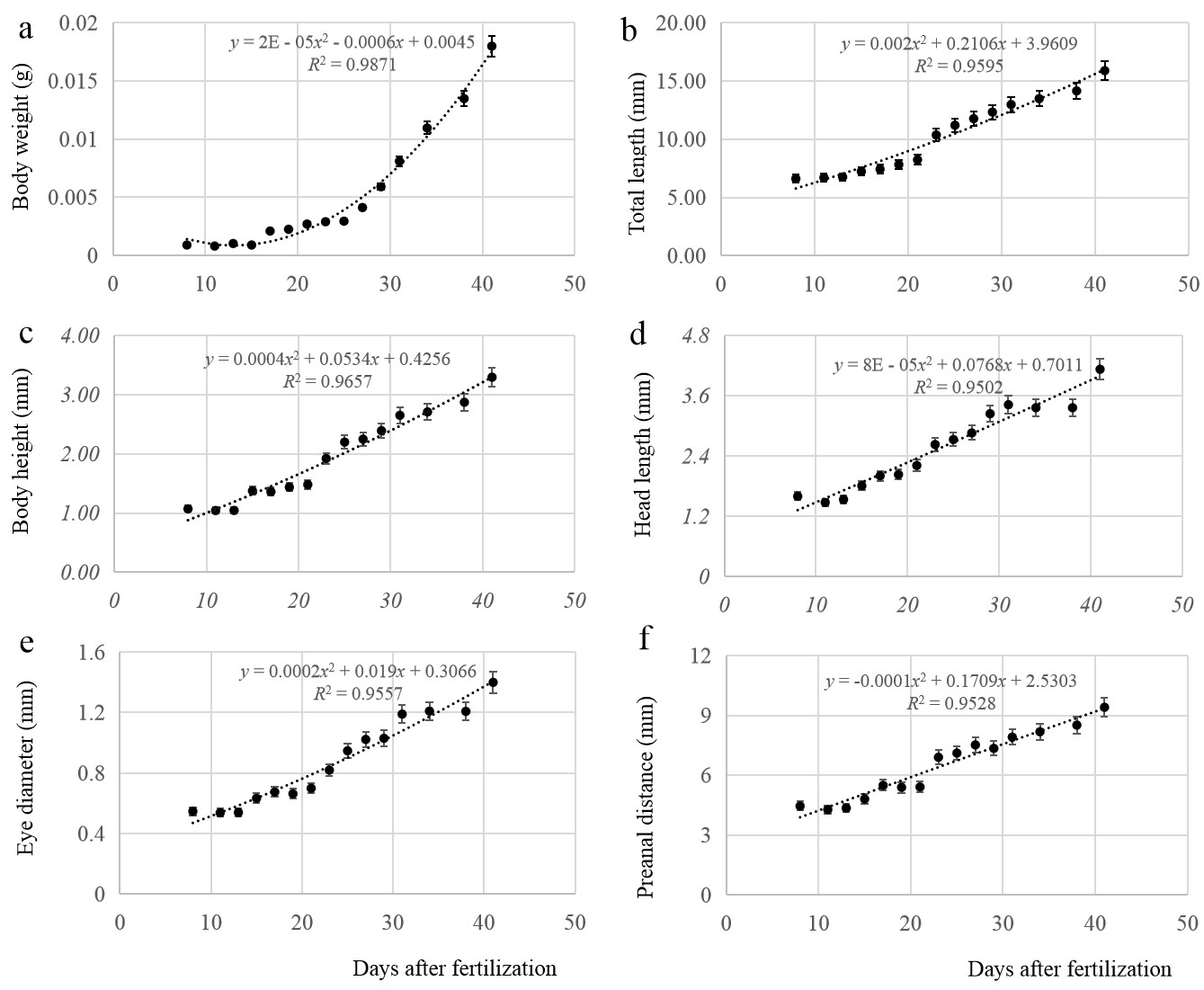
The relationships between body weight, total length, body height, head length, eye diameter, preanal distance, and time were fitted by regression equations. The models for body weight (Figure 2 a), total length (Figure 2 b), body height (Figure 2 c), head length (Figure 2 d), eye diameter (Figure 2 e), and preanal distance (Figure 2 f) among larvae can be described by the respective equations:
y=2E−05x2−0.0006x+0.0045,R2=0.9871
y=0.002x2+0.2106x+3.9609,R2=0.9595
y=0.0004x2+0.0534x+0.4256,R2=0.9657
y=8E−05x2+0.0768x+0.7011,R2=0.9502
y=0.0002x2+0.019x+0.3066,R2=0.9557
y=−0.0001x2+0.1709x+2.5303,R2=0.9528
In the above equations, y represents the growth parameter x is age (days), where x ∈ [8,41]. All parameters showed allometric growth during development, which was slow before 21 days of age and then accelerated.

Discussion
Early embryonic development of green crucian carp is generally consistent with that of other carp species, although each has unique characteristics. Chen et al.16 reported that it took 147 hr for the fertilized eggs of Aphyocypris lini to break the membrane at a temperature of 14–16.8 °C, 50 hr at 22.5–25 °C, and 28 hr at 28–30 °C. Therefore, the embryonic development rate is positively related to water temperature within a certain range. The embryos of green crucian carp broke the membrane after 56.5 hr at a water temperature of 24 ± 1 °C. Under similar temperatures, the embryonic development rate of green crucian carp had no obvious differences from those of other carp fishes. Poikilotherms must absorb heat from the environment during embryonic development, and the total heat needed in different development stages is the effective accumulative temperature. The effective accumulative temperature needed for green crucian carp to go from fertilization to hatching was 1361.7 °C x hr, which is far lower than required for Dianchi crucian carp (2413.3 °C x hr); however, it is quite close to that required for goldfish (1408 °C x hr).
The mature egg diameter of green crucian carp was 1.47 ± 0.04 mm, smaller than other related fishes. For example, the mature egg diameter of Carassius auratus pengzesis is about 1.8 mm, while the mature egg diameters of triploid crucian carp and Prussian carp are about 1.6 mm.12,19,20 Due to the small egg diameter, the egg number is increased, which increases the reproduction ability of green crucian carp to some extent. This might be consistent with their living environment. Yolk fluctuation phenomena have been observed in the multi-cell phase of embryonic development in some cyprinid fishes, such as Prussian carp and triploid crucian carp. However, such phenomena have not been observed in other cyprinid fishes, such as zebrafish23 and grass goldfish.26 The sarcomere is the basic unit muscles need to execute their stretching function. The sarcomere of green crucian carp was formed after the eye primordium’s development, similar to the triploid crucian carp’s.19 However, the sarcomere in Carassius auratus pengzesis is formed before the eye primordium.12 In the beginning, five pairs of sarcomeres were developed in green crucian carp, which is significantly higher than in Gaibei carp and Carassius auratus pengzesis (3–4 pairs). Surprisingly, the heart rate in the early hatching phase of green crucian carp was 136 beats/min, more than three times that of triploid crucian carp (39 beats/min19). This might be related to the early sarcomere development in green crucian carp.
Like in other bony fish, the total length, body height, head length, eye diameter, and preanal distance in green crucian carp larvae were positively correlated with time.27 The head develops in preference to total length and height and is important for vision, breathing and feeding, which are especially important in the early development stage. Eye diameter is an important indicator of the development of neurons and sensory organs in fish and directly affects the ability of juveniles to hunt. The head length and eye diameter developed slowly in the early incubation stage. The larvae began to show phototaxis and gradually showed scavenging and aggressive behaviors, possibly related to their living habits and insufficient breeding density.28 The early embryonic development of green crucian carp is adapted to the development of digestion, growth, and feeding to improve survival in the external environment. Therefore, it is of great significance to study the biological characteristics of green crucian carp in the early developmental stage to improve the survival rate of its hatchlings.
Conclusions
Existing studies on embryonic development in fish have mainly focused on common farmed fish species, with few focusing on regional endemic fish. As a new subspecies of crucian species in Dongting Lake, green crucian carp have many good traits, such as a rapid growth rate and high disease resistance. Studying the embryonic development of green crucian carp is of great significance. The timing and characteristics of different stages of embryonic development were determined in this study. In particular, the fertilized eggs of green crucian carp were small. Some unique characteristics, such as yolk fluctuation and accelerated heart rate before hatching, were observed during embryonic development. The results provide a reference for the breeding and cultivation of green crucian carp.
Ethical Statement
All animal experiments were conducted by the Guiding Principles for the Care and Use of Laboratory Animals and were approved by Hunan University of Arts and Science.
Funding information
The present study was supported by the National Natural Science Foundation of China (grant no. 32172965), the Hunan Natural Science Foundation (2022JJ30033), and the Hunan Provincial Education Department Key Fund (18A362).
Conflict of interest
The authors declare that they have no competing interests.
Acknowledgments
We thank Mr. Liye Shao for raising the green crucian carp used in this study. We also thank the staff of the Hunan Provincial Key Laboratory for Molecular Immunity Technology of Aquatic Animals for their contributions and technical assistance during the experiments.
Authors’ Contribution per CRediT
Conceptualization: Yunsheng Zhang (Equal), Hu Xia (Equal), Pinhong Yang (Equal). Data curation: Yunsheng Zhang (Equal), Sigang Li (Equal). Funding acquisition: Yunsheng Zhang (Lead). Investigation: Yunsheng Zhang (Equal), Sigang Li (Equal), Jiaqi Liu (Equal). Methodology: Yunsheng Zhang (Equal), Hu Xia (Equal). Project administration: Yunsheng Zhang (Equal), Hu Xia (Equal), Liangguo Liu (Equal). Resources: Yunsheng Zhang (Equal), Pinhong Yang (Equal). Software: Yunsheng Zhang (Equal), Hu Xia (Equal), Sigang Li (Equal). Supervision: Yunsheng Zhang (Equal), Liangguo Liu (Equal), Pinhong Yang (Equal). Validation: Yunsheng Zhang (Equal), Sigang Li (Equal), Jiaqi Liu (Equal). Writing – original draft: Yunsheng Zhang (Equal). Writing – review & editing: Yunsheng Zhang (Equal), Hu Xia (Equal). Formal Analysis: Hu Xia (Equal), Sigang Li (Equal). Visualization: Sigang Li (Equal), Jiaqi Liu (Equal).