Introduction
Chinese aquaculture industry accounts for the largest share of the world’s fishery production and is becoming increasingly important in providing a principal source of protein for the surging population.1,2 Groupers are a very diverse family of predatory fish with high commercial value and widely distributed throughout the tropical and subtropical seas in the world.3 Giant grouper (Epinephelus lanceolatus) is the largest species of grouper, with a maximum weight up to 400 kg.4 Tiger grouper (E. fuscoguttatus) is a popular mariculture species in China because of its fast growth rate.5 Hybridization of the two species (♀E. fuscoguttatus×♂E. lanceolatus) becomes one of the best candidates for aquaculture farmers in China due to the fast growth, favorable taste, and large size.
Vibrio harveyi is a marine fluorescent gram-negative bacterium,6,7 and is a core species of Vibrio that requires for sodium ions for growth and widely scattered in natural aquatic systems.8–11 V. harveyi is a pathogen causing vibriosis in extensive marine fish and shellfish and leading to the huge economic loss in the marine aquaculture industry.12,13 The increasing resistance to various chemotherapeutic drugs and drug residue become a direct threat to human health and safety.14 Vaccination has been shown to be one of the most effective alternatives in disease control strategies.15
Chitosan oligosaccharides (COS), the degraded products of chitosan, have been thoroughly proven as the versatile biological functions, including antioxidant, anti-coagulant, anti-inflammatory, anti-microbial, neuroprotective, and matrix metalloproteinases inhibitory effects.16 In addition, strong efficacy, low cost, and low side effects are also characteristics. Because of its abundant advantages, COS is widely applied as a feed additive and vaccine adjuvant.17 Some studies have shown that injecting COS or adding oral COS into diets effectively enhances the immune resistance against pathogens in teleost.18,19
Inactivated vaccines comprise killed pathogens losing pathogenicity and retaining antigenicity, activating a humoral immune response against the corresponding pathogens.20,21 Our previous study showed that COS as an adjuvant of the Vibrio harveyi formalin-killed cells (FKC) vaccine notably enhanced the immune protective effect of FKC.22,23 However, the mechanism of COS enhancing FKC effectiveness against V. harveyi in grouper is unclear. In this study, the grouper spleen vaccinated with FKC+COS or PBS was collected, and transcription was analyzed to clarify the change in the related-immune gene expression levels. The results enriched the theoretical basis for the molecular mechanism of COS enhancing the immune resistance of inactivated vaccines by further activating the fish’s immune system.
Materials and Methods
Fish
Healthy hybrid groupers (average body weight 30.0 ± 3.0 g) were purchased from the local commercial fish farm (Zhanjiang, Guangdong, China) and kept in an aerated tank filled with sand-filtered seawater of 25±1.0°C. Fish were fed with commercial diets twice daily and acclimatized for two weeks. All animal experiments were conducted in accordance with the ethical standards approved by the Guangdong Provincial Key Laboratory of Aquatic Animal Disease Control and Healthy Culture.
Preparation of FKC and adjuvant
V. harveyi strain ZJ0603 was isolated from the diseased grouper and preserved in our laboratory. FKC was prepared according to the method of Wei et al.23 in our previous study. Adjuvant COS was provided by Yuanye Bio-Technology Co., Ltd (Shanghai, China). The FKC was suspended at a concentration of 2.8 × 109 cfu/ml in a COS solution (FKC + COS) at 4 mg/ml.
Vaccination and total RNA extraction
The fish were randomly divided into 2 groups (40 fish per group). A group was injected intraperitoneally with 100 μl FKC + COS, and the other group was injected with 100 μl PBS. Two weeks later, the spleen tissue of fish was collected from the PBS group and FKC + COS group and stored at -80°C until use. Total RNA was extracted using. Total RNA was analyzed by agarose gel electrophoresis and Nanodrop (Bio-DL, Shanghai, China) to detect RNA purity (OD260/OD280 ratio). The Qubit system accurately quantified the RNA concentration, and the integrity of RNA was accurately detected by Agilent 2100 (Agilent Technologies). High-quality RNA samples were used for library construction.
Library preparation and sequencing Annotation and classification of genes
The RNA extraction libraries were constructed using PacBio single-molecule real-time (SMRT) technology. Novogene Technology Co., LTD (Guangzhou, China) performed high-quality, full-length transcript sequencing. In DEGs, 10 immune-related genes were selected for quantitative real-time PCR (qRT-PCR) analysis to verify the reliability of the results of RNA-seq analysis.
Annotation and classification of genes Quantitative real-time PCR validation
After sequencing, the raw data are corrected and de-redundantly processed to obtain high-quality data. To obtain functional annotations, non-redundant sequences were annotated in NR (NCBI non-redundant protein sequences), NT (NCBI nucleotide sequences), SwissProt (A manually annotated and reviewed protein sequence database), KEGG (Kyoto Encyclopedia of Genes and Genomes), KOG (Clusters of Orthologous Groups of proteins), GO (Gene Ontology) and Pfam (Protein family) databases. All differentially expressed genes (DEGs) were enriched and analyzed in GO (Gene Ontology: http://www.geneontology.org/) and KEGG (Kyoto Encyclopedia of Genes and Genomes: http://www.genome.jp/kegg/).
Quantitative real-time PCR validation
The qRT-PCR was carried out using TB Green® Premix Ex Taq™ II (Takara) in LightCycler 96® Real-Time System (Roche, CH). All data were analyzed with β-actin as an internal reference by the 2-ΔΔCt method. The reaction was performed under the following conditions: 95 °C for 5 min, 95 °C for 30 s,60 °C for 30 s and 72 °C for 30 s for 40 cycles. All primers were designed according to the corresponding sequences and are listed in Table 1.
Results
Transcriptional sequences annotation
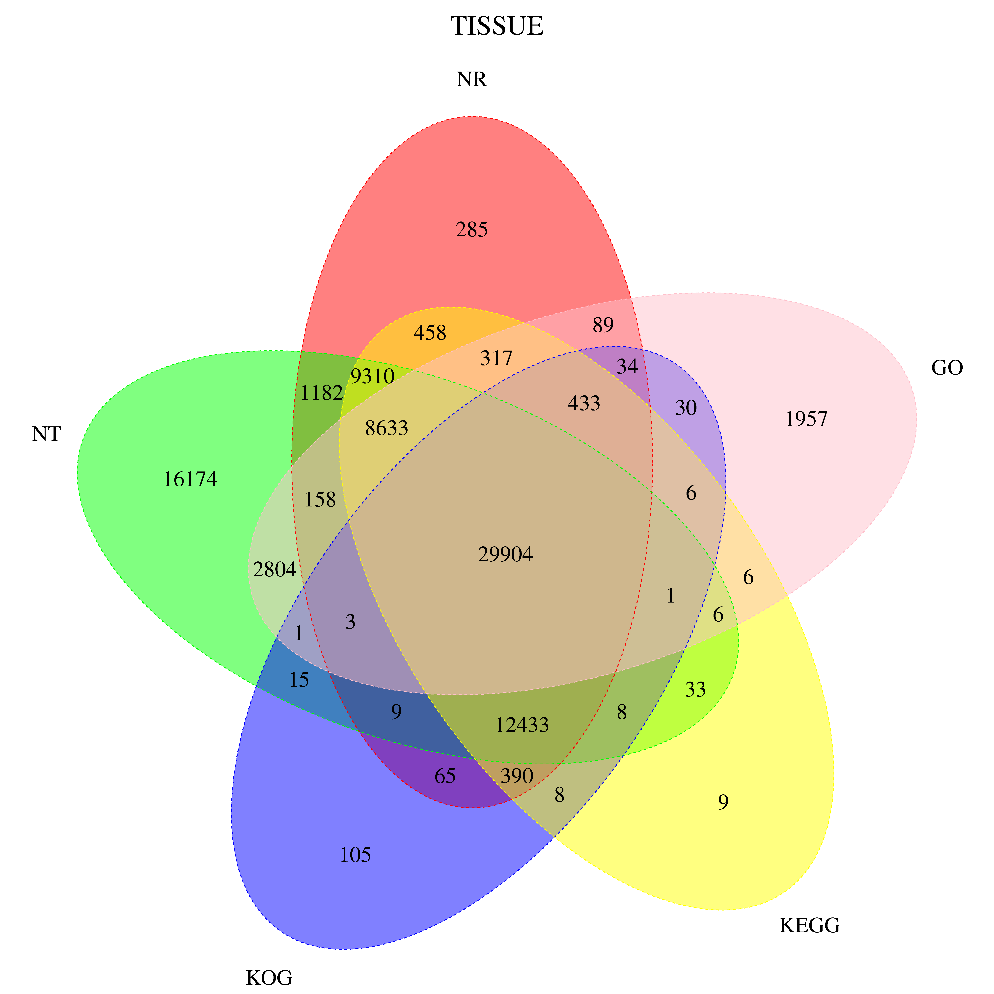
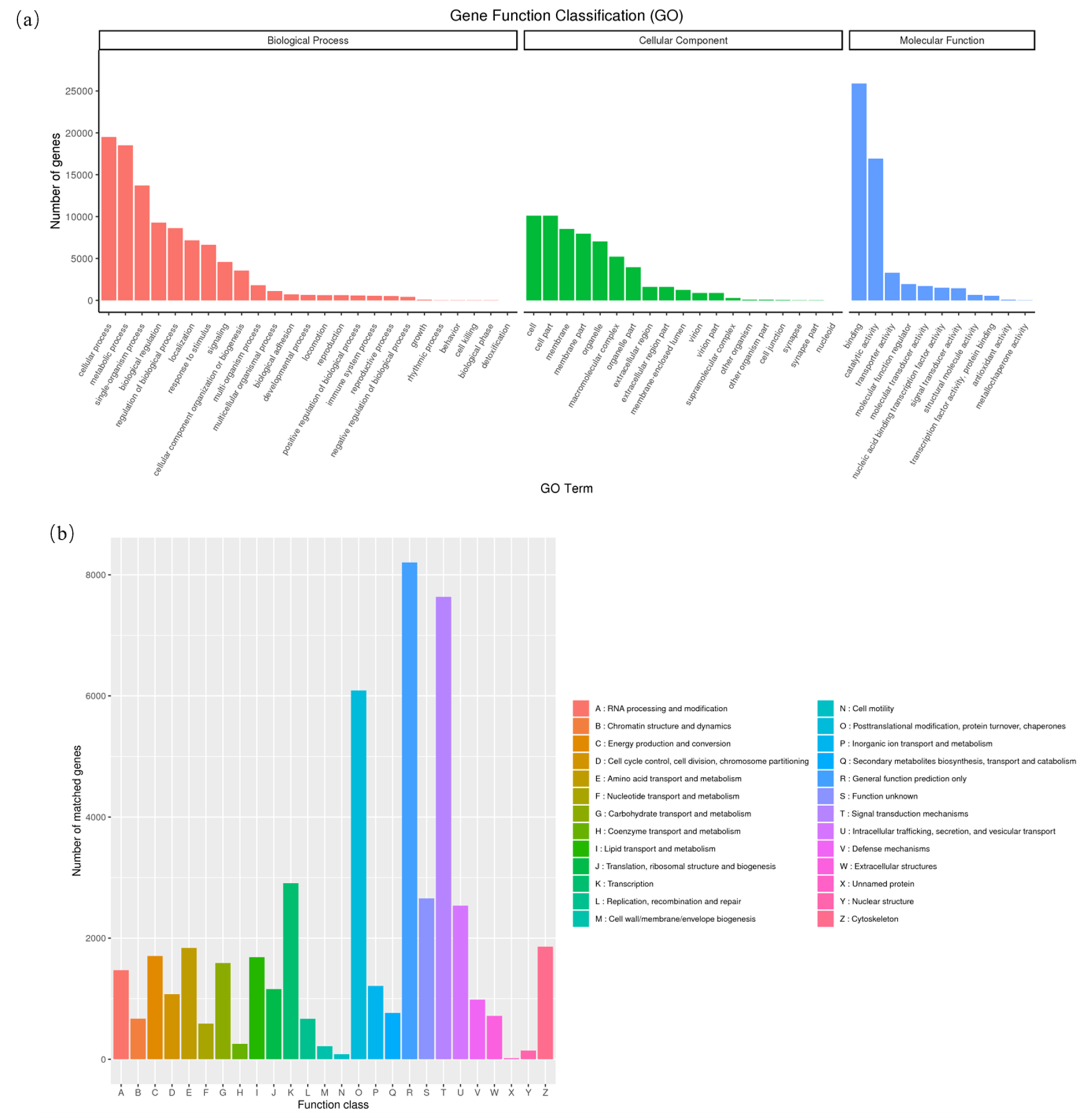
Transcriptome sequencing of the samples was performed (Figure 1 and Figure 2), and 47,714,744 and 50,615,040 raw reads were obtained in the samples injected with PBS and KFC+COS. To obtain gene function information, 236,341 non-redundant transcripts were annotated in 7 databases: 63,703 unigens in NR, 57,150 unigens in SwissProt, 61,955 unigens in KEGG, 43,445 unigens in KOG, 44,382 unigens in GO, 80,674 in unigens in NT, 44,382 unigens in Pfam and 29,865 unigens in all 7 databases. The 44,382 unigens annotated in GO (Gene Ontology) were classified into 3 categories: molecular function, biological process, and cellular component. The 43,445 unigens were classified into 26 categories in KOG, and 8,204 unigens belonged to R (General function prediction only), 7636 unigens belonged to T (Signal transduction mechanisms), 6090 unigens belonged to O (Posttranslational modification), and 17 unigens were classified as X (Unnamed).

_go_statistical_diagram_of_unigens__(.png)
DEGs analysis
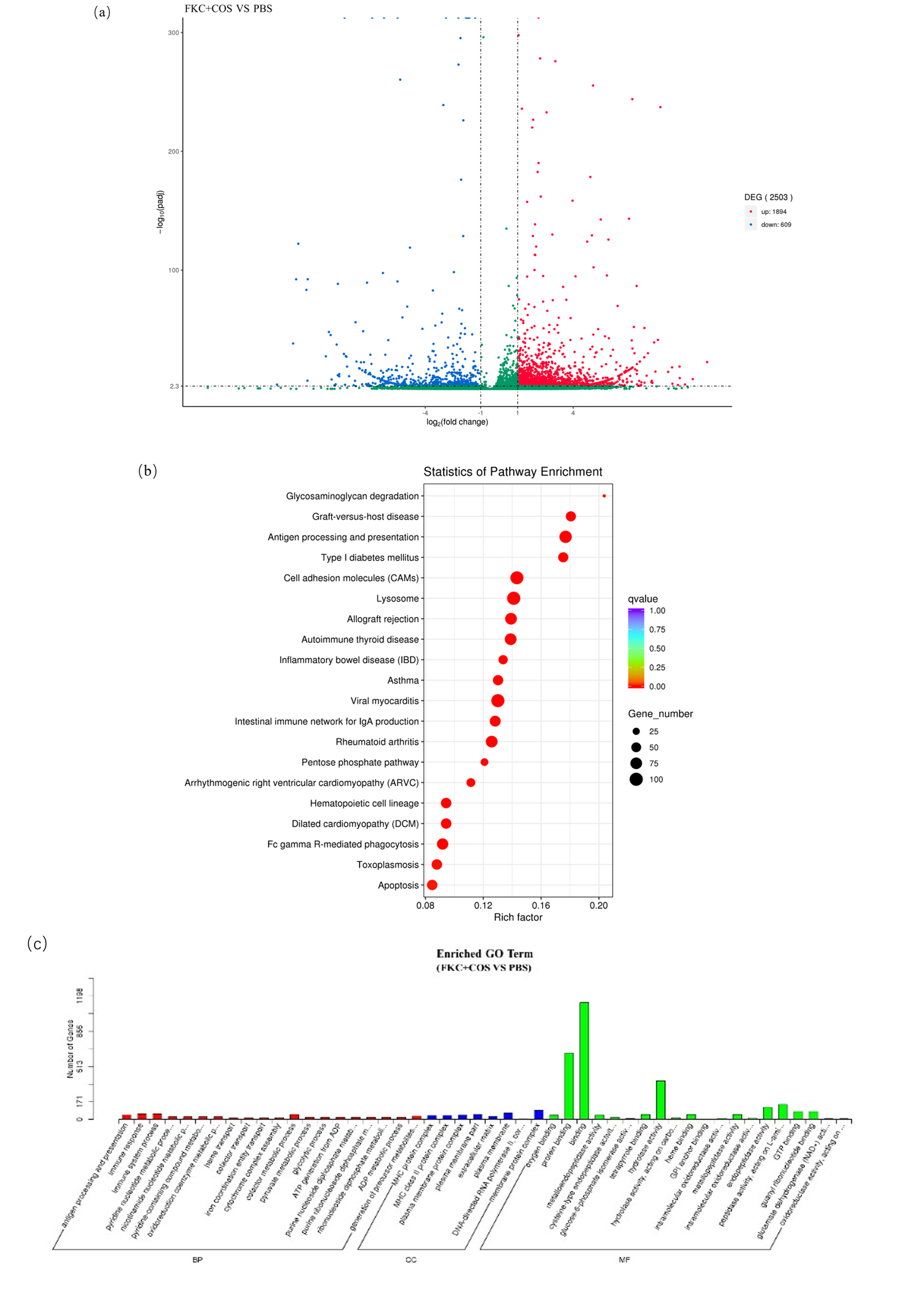
To further understand the effect of FKC+COS on the immune response of grouper, we analyzed DEGs of transcriptome sequencing data using FDR < 0.05 and |log2 (fold-change)| >2 as screening conditions (Figure 3). The results showed 2,503 unigenes with 1,894 upregulated genes and 609 downregulated genes. All DEGs were annotated in GO and KEGG for the function analysis. The three ontologies are classified into the biological process (BP), molecular function (MF), and cellular component (CC) according to GO annotation. GO database was further assigned to 48 functional terms, with 20 in BP, 20 in MF, and 8 in CC (Figure 3c). In KEGG analysis, the top 20 enriched pathways, the fish immune-related ones, were Antigen processing and presentation, Lysosome, Intestinal immune network for IgA production, and FcγR-mediated phagocytosis.
_volcano_map_for_differentially_expressed_gene.png)
Validation of results by qRT-PCR
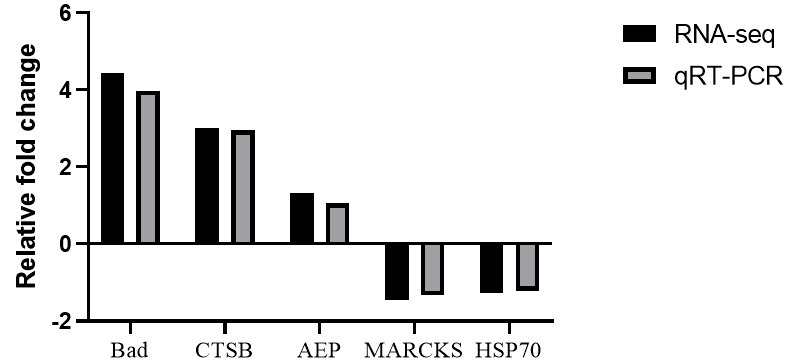
The qRT-PCR analysis was performed to confirm the RNA-seq data. The expression levels of 5 selected genes (Bad, CTSB, AEP, MARCKS, and HSP70) were measured and normalized to the expression of β-actin. As shown in Figure 4, the expression levels of 5 genes analyzed by qRT-PCR were mainly in agreement with the data of RNA-seq. The qRT-PCR analysis results confirmed that the data of RNA-seq were reliable.

Discussion
Of the superior palatability and nutritional value, hybrid grouper (♀ E. fuscoguttatus × ♂ E. lanceolatus) has become a very popular fish species among farmers and consumers.24 V. harveyi, which has been proven to be a highly pathogenic pathogen in marine fish, frequently threatens the health of grouper under the synergistic effect of an over-dense farming environment.25 Antibiotics and parasitic agents may be very effective, but their resistance and environmental damage are not tolerated. Effective vaccines have been the way to solve the disease problem, and many highly successful vaccines have been developed for aquaculture.26 COS is water-soluble, non-cytotoxic, well-biocompatible, and easily absorbed by the intestine.27 It has been shown to activate the immune system against pathogens, making it an attractive immune booster.14 Our previous study showed that the immune protection rate of a single FKC vaccine against grouper was 52%, while the immune protection rate of an FKC+COS combined vaccine against grouper was 80%. This proves that COS is an efficient immunostimulant to enhance the FKC effectiveness against V. harveyi in grouper against V. harveyi.23 However, the mechanism of COS enhancing the immune protective effect of FKC against vibriosis is unclear. To explore the role of COS as an adjuvant of FKC, we performed transcriptome analysis with grouper spleens tissue 14 days post-immunization of PBS and FKC+COS, respectively.
It is evident that understanding the immune response of grouper after vaccination is of great significance for selecting the vaccine strategy for grouper. We performed transcriptome sequencing of the spleen of the grouper, which was injected with FKC+COS and PBS. The 2,503 DEGs were obtained, including 2153 upregulated and 616 downregulated genes. KEGG analysis of DEGs found that the pathways highly related to immunity were antigen processing and presentation, lysosome, and intestinal immune network for IgA production and FcγR-mediated phagocytosis among the top 20 pathways with the highest enrichment. These results suggested that COS as an adjuvant of FKC could further enhance the grouper’s innate and adaptive immune.
Antigen processing and presentation are indispensable for adaptive immunity in teleost fish. Studies have shown that the functions of major histocompatibility complex II (MHC II) like molecules in this pathway in fish were similar to those in mammals.28–30 MHC II could interact with CD4+ T lymphocytes31,32 when antigen-presenting cells present motifs of extracellular danger signal from pathogens to specialized T cells (CD4±type) the MHC II mediated immunity is activated.33 In this study, the expressions of AEP and CTSB in MHC II are significantly upregulated. Asparaginyl Endopeptidase (AEP) is a lysosomal cysteine protease conserved in diverse cell types.34 it is involved in antigen presentation within MHC II positive cells and pro-protein processing.35 Cathepsin B (CTSB) has exopeptidase and endopeptidase activities and plays a vital role in various physiological degradation, apoptosis, and inflammatory response of many diseases.36 These results indicate that MHC II is activated in the grouper vaccinated with the combination vaccine. Similar responses also occurred in bacterial stress on Verasper variegates, Nile tilapia,w, and Paralichthys olivaceus.37–39 Active MHC II positively affects the recognition and clearance of pathogenic bacteria.
The terminology of FcγR-mediated phagocytosis mediating phagocytosis in KEGG is extremely important. Phagocytosis is an evolutionarily conserved process utilized by many cells to ingest microbial pathogens and apoptotic and necrotic corpses.40,41
Phagocytosis by macrophages can be initiated by the Fcγ receptors (FcR) in membranes which combine with Fc regions of immunoglobulin G (IgG).42 FcγR may also participate in the phagocytosis of mIgM lymphocytes.43
Notably, the expression of some lysosomal genes was significantly upregulated. Lysosomes are essential and well-equipped subcellular organelles in animals’ immune response, and contain a mass of hydrolases. Pathogenic microorganisms can be transported to lysosomes to be killed and degraded by various acids after autophagy or endocytosis.44,45 In addition to mediating the elimination of pathogens and harmful substances, lysosomes can still efficiently repair damaged plasma membranes during bacterial infections.46,47
This research also found that inflammatory bowel disease (IBD) expression levels dramatically increased. The NOD2, which was found to be significantly upregulated in IBD, is the founding member of the intracellular NOD-like receptor family and is mainly expressed by two cell types that are exposed to this component under physiological conditions: antigen-presenting cells (APCs) and epithelial cells.48 As a microbial sensor, NOD2 proteins operate could recognition of specific PG (peptidoglycan) constituents of bacteria and significantly activate NF-κB, IFNβ, and MAPK signaling pathways.49–51 The decreased NOD2 function could promote the invasion of bacteria and eventually lead to chronic intestinal inflammation. Its high expression in this study suggests that hybrid grouper NOD2 may play an essential part in the innate immune system.52
In summary, we analyzed the transcriptomic response of grouper vaccinated with FKC+COS and PBS based on high-throughput sequencing. We found some genes in the adaptive system associated with phagocytosis-associated antigen processing and presentation. FcγR-mediated phagocytosis mediating phagocytosis was significantly upregulated in the FKC+COS group versus the PBS group. Furthermore, the significantly high expression levels of some genes in an innate system associated with phagocytosis, lysosomes, and IBD were found in the FKC+COS group versus the PBS group. These results provide clues to explore the potential mechanism of COS as an adjuvant enhancing the protection effectiveness of FKC against vibriosis in grouper.
Declaration of competing interest
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
This work was funded by the Natural Science Foundation of Shenzhen City (JCYJ20190813114409506 and JCYJ20210324130003009) and the Natural Science Foundation of Guangdong Province (No. 2021A1515010532).