Introduction
Procambarus clarkii is a freshwater crayfish from Arthropoda, Crustacea, Decapoda, Reptantia, Cambaridae, Procambarus.1 It is native to north-eastern Mexico and the south-central United States2 and was introduced to China in 1920s. As one of the most invasive species, P. clarkii can adapt to almost all types of freshwater habitats, such as wetlands, marshes, small natural waterways, and paddy fields, and be found in most areas of China except the Tibet plateau.3,4 In recent years, P. clarkii has been extensively treated as a delicacy and consumed in large quantities in China, making it one of the most economically important farmed freshwater species.4
Estradiol is a steroid estrogen closely related to animal ovarian development. In vertebrates, estradiol induces vitellogenesis under the anterior pituitary and hypothalamus regulation, thereby promoting ovarian development. In recent years, more and more studies have shown that invertebrates can also secrete estrogen that promotes ovarian development. There is evidence to prove that the ovaries, hepatopancreas, and blood of shrimp and crabs contain a certain concentration of estradiol, and its content is related to ovarian development.5 Extracorporeal injection of estradiol can increase oocyte diameter and gonadal index, promoting ovarian development.6,7
Moreover, estrogen synthesis-related enzymes are found in crustaceans. In the hepatopancreas and ovary of Marsupenaeus japonicus, 17-hydroxysteroid dehydrogenase, a key enzyme in steroid hormone metabolism, is detected. The activity of this enzyme in the hepatopancreas of mature female shrimp is significantly higher than that in immature female shrimp. In addition, estradiol can increase the activity of this enzyme.7 This indicates that estradiol plays an important role in the development of crustacean ovaries.
The hepatopancreas is the main organ for lipid storage and processing in crustaceans, playing a crucial role in their growth, development, and reproduction. Crustacean ovaries’ protein and fat contents significantly increase during development and maturation. The increase in protein and fat in the ovaries is related to the corresponding decrease in these substances in the hepatopancreas. During the ovarian development of decapod crustaceans, the hepatopancreas continuously synthesizes vitellogenin, lipids, and other substances and transports them to the developing oocytes through hemolymph, providing energy for the development of oocytes.5,8,9
The type and content of fatty acids in the ovary of P. clarkii undergo significant changes during its development, which will naturally be reflected in the hepatopancreas, the main organ that provides nutrition for ovarian development.10 Previous studies found that estradiol not only existed in the hepatopancreas of P. clarkii, but was also related to ovarian development.10 Thus, this study aimed to investigate the effect of exogenous estradiol injection on the fatty acid content and tissue structure of the hepatopancreas of P. clarkii, to provide a reference for studying the roles of estradiol in ovarian development in crustaceans.
Materials and Methods
Experimental crayfish and its feeding management
The female parent individuals of P. clarkii were purchased from Renmin Road Market in Yancheng City. The selected individuals have strong vitality, large size, red body color, no disease or injury, and strong abdominal limbs. Total number: 240 individuals; Specification: total body length is (9.82±0.55) cm, length of the carapace is (4.46±0.30) cm, and body weight is (20.43±2.45) g. The female crayfish were taken back to the laboratory and temporarily raised in square aquariums (60 cm×45 cm×50 cm) with a water depth of 15cm. 5 arched tiles were placed in each aquarium to prevent crayfish killing. Continuous aeration of water was performed. The crayfish were fed with fresh Unionidae weighing 5%-7% of the body weight of the crayfish daily. 25% water was changed to maintain water refreshment.
Injection of estradiol
The crayfish were injected with estradiol of different concentrations. The experiment was designed as four groups: (1) control group, physiological saline injection; (2) 0.05 group, 0.05 µg/(g body weight) estradiol injection; 0.5 group, 0.5 µg/(g body weight) estradiol injection, and 5 µg/g group, 5 µg/(g body weight) estradiol injection. The injection volume is 100 µL/individual. The specific process is to hold the crayfish with the left hand to prevent it from squirming free, hold the syringe with the right hand, inhale the prepared medicine, and slowly inject it into the crayfish muscle between the second and third abdominal shells at a 45° angle to the body. Due to the hard shell of the lobster shell, the operation was carefully conducted to ensure not to put the syringe on the abdominal shell. The crayfish in the control group were injected with 100 µL/individual physiological saline. Each treatment was repeated for three times, and each duplicate contained 20 individuals.
The injection was conducted once every 5 days, a total of 5 times. The whole experiment lasted 25 days. The same injection site was not used for the adjacent two injections to reduce harm to the crayfish. The injection dates are October 23rd, October 28th, November 2nd, November 7th, and November 12th in 2022. After the last injection, the crayfish were temporarily raised for 24 hours. The hepatopancreas samples were taken on November 13th, and stored at -72°C for further determination.
Preparation of tissue slices
Paraffin slice preparation: (1) First, the fixed samples were subjected to gradient dehydration with alcohol gradients of 70%, 80%, 95%, and 100%; (2) then, the samples were made to be transparent with ethanol xylene and xylene I and xylene II in a volume ratio of 1:1:1; (3) finally, the samples were embedded in paraffin, sliced with Leica slicer with a thickness of 6 μm. Slices were stained with H.E contrast, then observed and taken photos using a Nikon80i research-grade microscope.
Determination of fatty acids
Referring to the Folch method,11 We weighed approximately 0.4 g of the hepatopancreas sample, placed it in a stoppered test tube, and added 10 mL of chloroform-methanol solution (2:1, v/v). The tubes were shaken in the oscillator for 5 minutes, then the lipids were extracted under a sealed and dark condition for 8 hours. After extraction, 2 mL of 0.9% NaCl aqueous solution was added to each tube, shaken for 30 seconds, and then let it stand for layering. A pipette was used to suck 6 mL of chloroform in the lower layer into a 10 mL centrifuge tube, then 1 mL of 0.9% NaCl aqueous solution was added, followed by shaking again for 30 seconds and then put the tubes into a centrifuge for centrifugation for 5 minutes (2000r/min). After centrifugation, 5 mL of the lower layer of chloroform was sucked and transferred to another centrifuge tube, then using the same method to centrifuge the tubes again. After secondary centrifugation, 3 mL of the lower layer of chloroform was sucked, and the solvent was removed from samples through rotary evaporation to obtain lipids.
After extraction with 1 mL heptane, the extracts were placed in a 10 mL tube with a stopper. Then 1 mL of 1mol/L sodium hydroxide-methanol solution was added into the tube, and then the tube was placed in a 70 oC water bath for esterification for 30 minutes. After esterification, 2 mL heptane was added to the test tube, followed by 2 mL of saturated NaCl aqueous solution addition, and let it stand for layering. A pipette was used to absorb 1 mL of the upper heptane and put it into a 1.5 mL centrifuge tube. About 0.3 g of anhydrous sodium sulfate was added to the centrifuge tube. Then centrifuged for 5 minutes (1×1000r/min). The centrifuged fatty acid methyl ester was stored at 3oC for future analysis.
1 µL fatty acid methyl ester sample was sucked for gas chromatography-mass spectrometry analysis. The analysis conditions were: analytical instrument: Trace DSQ GC/MS gas chromatography-mass spectrometry; Chromatographic column: HP-5MS, 30 m×0.25 mm×0.25 μm. Gas chromatography operating conditions: The gasification chamber temperature was set at 250 oC, and the transmission line temperature was set at 280 oC. The temperature rises process of the chromatographic column: initial temperature 50 oC, increase to 280 oC at 10 oC/minute, and maintained for 10 minutes. Injection method: split injection, with a split ratio of 10:1. Injection volume: 1 μL. Mass spectrometry: EI ion source, signal amplifier voltage: 1200 V. Ion source temperature: 230 oC. Quadrupole temperature: 150 oC, full scan (SCAN) quality range: 45-500 mau. The type of fatty acids in the sample can be determined by retrieving the NIST mass spectrometry library and comparing the sample mass spectrometry with the standard mass spectrometry in the library. The relative content of each fatty acid was calculated using the area normalization method.
Results
Effects of estradiol on fatty acid content in the hepatopancreas of P. clarkii
The main saturated fatty acids (SFA) in the hepatopancreas of P. clarkii are C18:0 and C16:0. The main monounsaturated fatty acid (MUFA) is C18:1n9. And the main polyunsaturated fatty acids (PUFA) are C18:4n and C20:3n (Table 1).
After 25 days, injecting estradiol significantly reduced the total content of PUFA in hepatopancreas (P<0.05), and significantly increased the total content of SFA and MUFA (P<0.05). Specifically, injecting estradiol significantly increased C16:0, C20:1n, C20:2n, C22:n (n=1-4) and DHA (P<0.05). Injecting estradiol significantly reduced C18:4n, C20:3n, EPA and C22:n (n=1-4) (P<0.05). Injecting 5 and 0.5 μg/g estradiol significantly increased C18:1n9 and C18:2n (P<0.05). Injecting 0.5 and 0.05 μg/g estradiol significantly reduced C18:3n6 (P<0.05). Injecting 0.05 μg/g estradiol significantly increased C16:1n, C18:0 and C22:n (n=1-4), and significantly reduced C20:4n3 and DPA (P<0.05, Table 1).
Effects of estradiol on the tissue structure of the hepatopancreas of P. clarkii
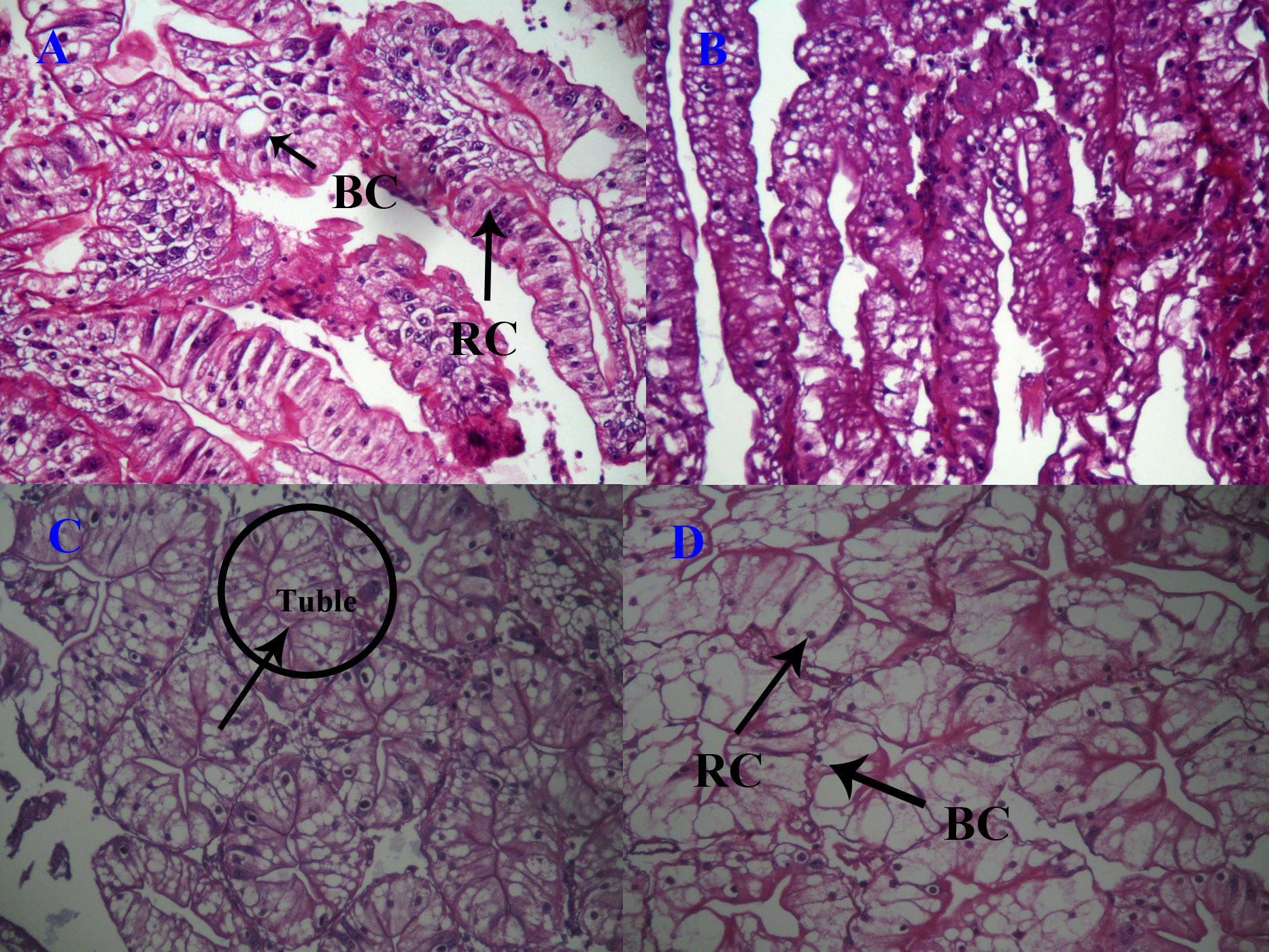
Blasenzellen cells (B cells), fibrillenzellen cells (F cells), restzellen cells (R cells), and embryonalzellen cells (E cells) can be observed in the hepatopancreas of P. clarkii. B and R cells were dominant, while the number of F and E cells was relatively small. The control group had smaller liver corpuscles of hepatopancreas, and they were arranged densely, with a more uniform distribution of B and R cells (Figure 1A). After injecting 0.05 μg/g estradiol, the number and volume of B cells increased, and vacuoles in B cells increased. The number of R cells showed no obvious difference compared to the control group (Figure 1B). Injecting 0.5 μg/g estradiol further increased the volume and vacuoles of B cells, while the number of R cells decreased (Figure 1C). After injecting 5 μg/g estradiol, cell volume and vacuoles of B cells continued to increase, while the number of R cells further decreased (Figure 1D).
.jpeg)
Discussion
Effects of estradiol on the content of fatty acids in the hepatopancreas of P. clarkii
The determination of this experiment showed that the top three fatty acids in the hepatopancreas of P. clarkii were C18:4n (parinaric acid), C20:3n (eicosatrienoic acid), and EPA, with the highest content of eicosatrienoic acid. The results are different from previous studies, which commonly found that C18:n9 (oleic acid) and C18:2n6 (linoleic acid) were the main fatty acids in the hepatopancreas of P. clarkii.10,12,13 The hepatopancreas is the main organ for fat storage in crustaceans, and the changes in fatty acid content are closely related to the changes in fatty acid content during ovarian development. During the maturation process of ovaries, the content of certain fatty acids increases while the corresponding fatty acids decrease in the hepatopancreas (Amran et al., 2018). C18:n9 (oleic acid) and C18:2n6 (linoleic acid) were found in crustaceans and ovaries, and when the oleic acid and linoleic acid were consumed in crustaceans, they accumulated in ovaries. The phenomenon was found in many Decapoda crustaceans,9,14,15 which proved that the oleic acid and linoleic acid in crustaceans were the energy supply substance for ovarian development.
Interestingly, in this study, estradiol injection significantly increased oleic acid and linoleic acid content in P. clarkii, making oleic acid the highest fatty acid content. Compared to previous studies, the crayfish we used for this experiment were larger, which might have more developed ovaries, and more oleic acid and linoleic acid were consumed in crustaceans. Estradiol injection probably induced the oleic acid and linoleic acid accumulation in the hepatopancreas for preparation for ovarian development. It was also proved by the increase of C16:0 (palmitic acid) induced by estradiol injection, which might be connected to ovarian development. To better understand the process, the dynamics of fatty acids components at different development stages of P. clarkii and the mechanisms need to study in the future.
Effects of estradiol on the tissue structure of the hepatopancreas of P. clarkii
The hepatopancreas are the main digestive organs of crustaceans, responsible for many important physiological functions, and changes in the tissue structure reflect changes in the physiological state of the body. Like most crustaceans,16 the hepatopancreas of P. clarkii consists of four types of cells, namely, B cells, F cells, R cells and E cells. The main function of B cells is to secrete digestive enzymes; the function of F cells is to absorb nutrients; the function of R cells is to store nutrients transported from absorbing cells; and the function of E cells have differentiation potential and the ability to maintain the minimum survival. This study found that injection of estradiol led to an increase in vacuoles in the hepatopancreatic B cells of P. clarkii and a decrease in the number of R cells. Studies have shown that changes in the environment can cause changes in the proportion of cells in the hepatopancreas of crustaceans, especially B and R cells.17–20 R cells can store carbohydrates, lipids and ions (Verri et al., 2000). When the accumulation of substances mentioned above in R cells reaches a certain extent, the R cells change to B cells. Because B cells not only have the function of digestion and absorption but also can transport the accumulated substances to the lumen of proximal tubule.17–20 This can explain the phenomenon that estradiol injection increased the vacuoles in B cells and decreased the number of R cells. After estradiol injection, the content of certain fatty acids increased and stored in R cells, then R cells transported them to B cells for digestion and absorption, leading to the vacuole’s expansion in B cells. Sreeram and Menon21 found the proliferation of vacuolated B cell was almost up to the distal end of the tubules in the hepatopancreas of Metapenaeus dobsoni when exposed to 8 ppm petroleum hydrocarbons for 8 days. Naphthalene could also lead to the formation of large apical vacuoles in B cells of the hepatopancreas of Uca minax.22 When exposed to wastewater, B cell vacuoles in the hepatopancreas of Neosarmatium africanum became wider, indicating that the intracellular digestive activity increased through higher vacuolization processes and tubule dilation.23 In addition, 5 μg/g estradiol injection led to more R cell production in the hepatopancreas of P. clarkii than 0.05 and 0.5 μg/g estradiol injection, indicating that the effects of estradiol on the structure of hepatopancreas are related to the estradiol concentration. The higher concentration of estradiol injection led to an increase in the lipid content of hepatopancreas cells connected with the increased number of R cells.24 This study concluded that B and R cells play a crucial role in the metabolism and transportation of substances in the hepatopancreas of crustaceans.
Acknowledgments
This study was funded by Funding for school-level research projects of the Yancheng Institute of Technology (XJR2021035) and the Yancheng Key Research and Development Plan project (BE2023026).
Author contributions
Conceptualization: Weihong Zhao (Equal), Yanming Sui (Equal). Writing – original draft: Weihong Zhao (Equal), Liang Zheng (Equal). Funding acquisition: Weihong Zhao (Lead). Investigation: Jintao Liu (Equal), Jiyi Chen (Equal), Jiamei Wang (Equal). Methodology: Xirui Zheng (Lead), Liang Zheng (Equal). Writing – review & editing: Linlan Lv (Equal), Yanming Sui (Equal). Formal Analysis: Jiyi Chen (Lead), Jiamei Wang (Equal). Resources: Yanming Sui (Lead). Supervision: Liang Zheng (Lead).