


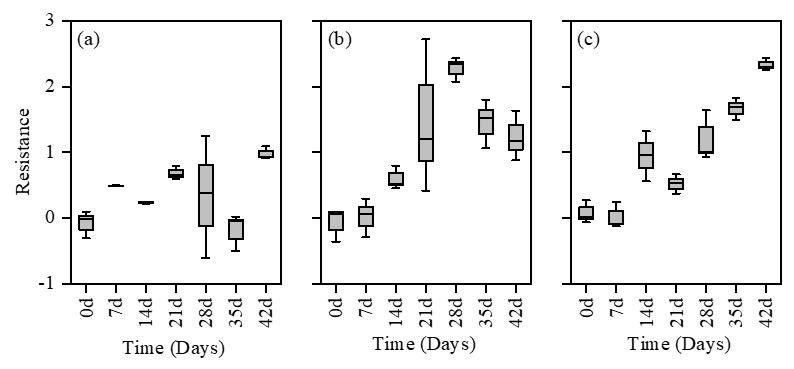
Introduction
Lakes are essential for maintaining the Earth’s water ecosystem, providing habitats for diverse organisms. However, they are highly vulnerable to the impacts of climate change and human activities.1 Phytoplankton, among the various organisms in lakes, holds immense significance. These microscopic organisms act as primary producers in freshwater environments, crucial in the food chain and material cycle. Additionally, they contribute to the flow of energy and transmission of information within the ecosystem.2,3 Different species of phytoplankton exhibit varying levels of sensitivity and adaptability to changes in lake environments. Therefore, assessing ecological parameters such as the structure and diversity of phytoplankton communities has become a common practice for evaluating the health of waterbodies.4,5
Human activities have significantly impacted climate change, leading to noticeable alterations in the frequency, severity, and timing of extreme weather events like heavy rainfall, typhoons, droughts, and heat waves.6 These events have substantially affected freshwater ecosystems, disrupting their functions and services. In recent years, unprecedented events such as the 2003 European heatwave,7 Australia’s “once-in-a-thousand-year drought”,8 and extreme rainfalls and high temperatures in Lake Erie, United States,9 have caused extensive damage to freshwater environments. These extreme weather events significantly drive change in freshwater ecosystems, impacting various levels of the aquatic ecosystem, including populations, communities, and ecosystems.10,11 Despite increasing research on the responses of biological communities to climate change over the past two decades, our understanding of the potential adaptation mechanisms for these communities in dealing with catastrophic events is still limited. This is primarily due to the severity of these extreme weather events and the challenges associated with conducting field studies in such conditions.12,13
In recent years, the Jianghan Plain has experienced a rise in heavy rainfall due to climate change and human activities. These events have caused flooding in and around Lake Changhu, damaging its ecosystems substantially. The lake has suffered from severe water pollution, a decline in aquatic plants and fish resources, and a deterioration of plankton structure.14,15 Additionally, the reduction in wetland areas has negatively impacted the biological function of the lake.16 With the expected increase in the frequency and intensity of extreme climate events, it is essential to understand the impact of high-frequency intense rainfall events on freshwater ecosystems.
Situated within the Jianghan Plain, Lake Changhu is a significant focal point known for its large size and shallow depth. Designated as a national aquatic resource reserve, it plays a critical role in maintaining the ecological balance in the middle reaches of the Yangtze River. The lake serves multiple purposes: flood control, water storage, agricultural irrigation, fish farming, shipping, and household water supply. However, the lake has faced several challenges in recent years. It has experienced a noticeable increase in the frequency of wet and rainy days during the spring and autumn seasons, accompanied by floods in early summer and heatwaves in the autumn season.17 Furthermore, previous studies have highlighted the impact of sewage discharge and enclosure on Lake Changhu, leading to eutrophication. This eutrophication has resulted in a decline in biodiversity across various trophic groups, including plankton, macrophytes, and fish.18–20
This research aims to explore the effects of varying rainfall patterns on the ecological status and phytoplankton community of Lake Changhu through in situ simulation experiments. Additionally, the study seeks to analyze the impact of terrestrial inputs induced by extreme rainfall on the composition and functionality of the phytoplankton community and evaluate its stability and ability to rebound when confronted with intense rainfall events. The findings of this study will provide valuable insights into aquatic ecosystems, emphasizing the importance of conserving robust freshwater ecosystems for the well-being of both humans and the environment.
Materials and methods
Study area
Lake Changhu (30°22’~30°30’ N, 112°17’~112°30’ E), located at the meeting point of Jingzhou, Jingmen, and Qianjiang cities, is the third largest freshwater lake in Hubei Province, China. Covering an area of approximately 122 to 150 km2 and boasting a shoreline that stretches about 310 km, this lake has a maximum water depth of 6.1 m and an average depth of 2.1 m, with a storage capacity of 2.7×108 m3. The region experiences a combination of rainfall and hot weather, with most rainfall occurring from April to August, coinciding with the hot weather season. During this time, the lake is primarily replenished by surface runoff and precipitation on its surface.
Lake Changhu is predominantly oriented east-west, with a broad eastern lake area and a narrower western section. The surrounding dykes and canals are essential for regulating water levels and reducing flood risks, especially during periods of heavy rainfall. The lake’s numerous bays and branches provide vital habitats for various aquatic plants and animals. The unique topography of Lake Changhu significantly contributes to its importance as a crucial ecosystem for biodiversity and the provision of essential ecosystem services.
Experimental design
The experiment was conducted at Lake Changhu Field Research Station of Yangtze University, situated on the west edge of Lake Changhu, Hubei Province, China. The experiment took place from 25th April to 15th June 2021. A one-factor experimental design was employed based on a comprehensive review of existing literature and field investigations. This design consisted of four levels of terrigenic treatments involving nutrient additions: CK (control, no nutrient addition), LT (low terrigenic inputs, with nitrogen and phosphorus concentrations 1.0 mg/L and 0.05 mg/L higher than CK, respectively), MT (medium terrigenic inputs, with nitrogen and phosphorus concentrations 2.0 mg/L and 0.1 mg/L higher than CK, respectively), and HT (high terrigenic inputs, with nitrogen and phosphorus concentrations 4.0 mg/L and 0.2 mg/L higher than CK, respectively). Each treatment level was replicated three times. To experiment, twelve square polypropylene bags (inner length: 1.0 m; width: 1.0 m; height: 1.0 m; volume: 1000 L) were filled with 800 L of water sourced from Lake Changhu. These bags were then suspended on the water surfaces, as illustrated in Figure 1. After allowing the water to stabilize for five days, allochthonous nutrients were added to the bags as part of the terrigenic treatment. Sodium bicarbonate (NaHCO3), potassium nitrate (KNO3), and dipotassium hydrogen phosphate (K2HPO4) were utilized to simulate the input of carbon, nitrogen, and phosphorus from terrestrial sources, respectively. To ensure proper hydrodynamics within the water column and create a habitat with low light levels, all mesocosms were mixed every three days and covered with shade nets. Additionally, the bags were shielded with Plexiglass that was transparent to ultraviolet radiation, thereby preventing the impact of precipitation on the experiment.
._there_were_three_replicate_.png)
Sampling and processing
Water samples from all mesocosms were collected from 6-day intervals from April to July 2021, typically between 8:00 and 10:00 a.m. The in-situ measurement of water temperature (WT) and dissolved oxygen (DO) was conducted using a Hydrolab DS5 multi-parameter water quality analyzer (Hach, Loveland, CO, USA). Samples were gathered using a 5-L polyethylene sampler for water chemistry and phytoplankton analysis. Phytoplankton samples were preserved in situ with 1.5% acid Lugol’s solution.
The collected water samples were promptly subjected to chemical analysis following established protocols.21 Standard methods were employed to determine the chemical characteristics of the water, including total nitrogen (TN), total dissolved nitrogen (TDN), ammonium (NH4+-N), nitrate (NO3--N), total phosphorus (TP), and total dissolved phosphorus (TDP). To conduct phytoplankton taxonomic enumeration, the collected samples were concentrated to a volume of 30 mL after allowing 48 h for sedimentation. Taxonomic identification of phytoplankton taxa was carried out at the species level under a microscope (Olympus CX23) at a magnification of 400×, following the methodology outlined by John et al.22 and Hu & Wei.23 For each phytoplankton sample, a minimum of 500 individuals were enumerated.24 The cell size of each counted individual was determined by measuring at least 30 cells of each taxon. Estimating species cell biovolume was accomplished using standard geometric formulae based on the recorded cell numbers and cell size measurements.25
Statistics and analysis
Differences across terrigenic treatments were tested using analysis of variance (ANOVA). The formula for the Shannon-Wiener diversity (H′), Pielou evenness (J), and Mcnaughton dominance (Y) indices are as follows:
H′=−s∑i=1PilnPi
J=H′/log2S
Y=niNfi
where S is the number of species, N is the total number of all algae, Pi is the proportion of the number of species i to the total number, ni is the number of species i, and fi is the frequency of occurrence of species i at each sampling stage.
Phytoplankton resource use efficiency (RUE) was quantified as phytoplankton biomass per unit total phosphorus.26 Resistance, on the other hand, was determined for the total abundance of phytoplankton by employing the approach outlined by Hillebrand et al.27 The difference between the perturbed and the control community was calculated using the log response ratio (LTR, resistance = ln (perturbed treatment/control treatment)) of the total phytoplankton abundance. Since the disturbance in our study persisted for over a month (extended pulse disturbance), resistance was calculated using data from all seven sampling events conducted throughout the experiment rather than just relying on data from the initial sampling event.
All statistical analyses were performed by using R software version 4.2.1 (R Development Core Team, 2022).
Results
Temporal variations in environmental factors
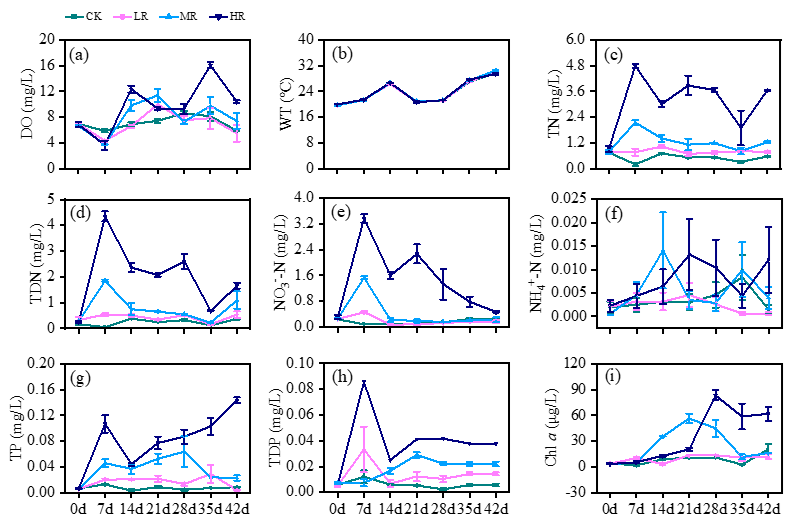
The environmental factors displayed consistent variation trends across the different treatments throughout the experiment (Figure 2). In the HT treatment, dissolved oxygen (DO) consistently increased from the beginning to the end of the experiment. Conversely, DO declined after three weeks in the CK, LT, and MT treatments (Figure 2a). Water temperature (WT) experienced a continuous increase throughout the experiment, with the highest temperatures recorded at 35 d in the MT treatment (30.47 ± 0.13 °C; Figure 2b). Total nitrogen (TN), total dissolved nitrogen (TDN), and nitrate nitrogen (NO3--N) demonstrated a decreasing trend from 7 d until the end of the experiment. The highest values recorded for TN, TDN, and NO3--N were 4.79 ± 0.09, 4.35 ± 0.19, and 3.37 ± 0.13 mg/L, respectively. Ammonium (NH4+-N) declined from 14 d until the end of the experiment, with the highest values observed at 0.014 ± 0.008 mg/L (Figure 2c-f). Total phosphorus (TP) and total dissolved phosphorus (TDP) displayed significant temporal fluctuations, showing variations among the different treatments (Figure 2g, h). Chlorophyll a (Chl a) demonstrated a continuous increase from the beginning to the end of the experiment in the CK and LR treatments. In contrast, in the MT and HT treatments, it decreased after the initial four weeks (Figure 2i).
_dissolved_oxygen_(do)__(b)_water_temperature_(wt)__(c)_total_nitrogen_(.png)
Phytoplankton dynamics
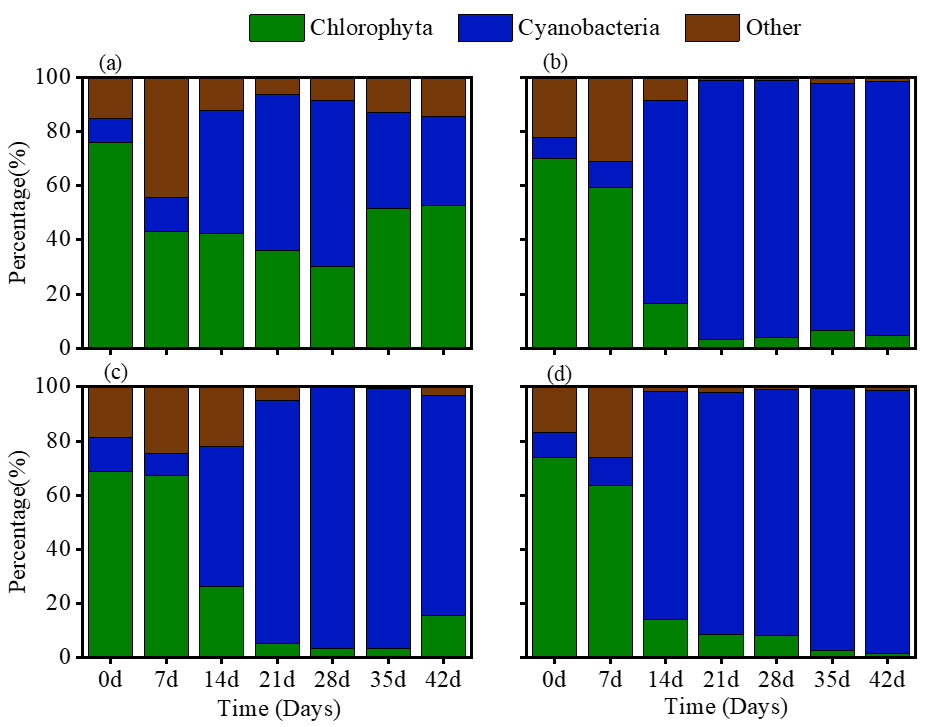
The phytoplankton community in all experiment treatments was primarily dominated by Chlorophyta and Cyanobacteria (Figure 3). In the CK treatment, Chlorophyta and Cyanobacteria accounted for approximately 40% of the total phytoplankton abundance (Figure 3a). However, in the treatments receiving terrigenic inputs, the Chlorophyta proportion significantly decreased over time (Figure 3b-d). Conversely, Cyanobacteria displayed an opposite trend to Chlorophyta, which contributed to more than 80% of the total phytoplankton abundance after the first week.

The introduction of terrigenic inputs had a detrimental impact on the species richness of phytoplankton (Figure 4a). The mean species richness decreased from 30.62 ± 3.90 in the CK treatment to 26.10 ± 4.06 in the HT treatment. Conversely, the total phytoplankton abundance exhibited a significant (P < 0.05) positive response to increased terrigenic inputs. The mean abundance increased from (0.91 ± 0.43) × 107 cells/L in the CK treatment to (19.18 ± 20.01) × 107 cells/L in the HT treatment (Figure 4b).
_and_total_phytoplankton_abundance_(b)_in_different_terr.png)
93 phytoplankton genera belonging to seven major taxonomic categories were identified throughout the experiment, with 11 dominant taxa identified (Table 1). In the CK treatment, the dominant phytoplankton species were primarily Chlorophyta (i.e. Chlorella sp. and Tetraedron regulare) and Cyanobacteria (i.e. Pseudanabaena sp., Aphanizomenon flosaquae, Anabaena sp., etc.). However, there was a noticeable shift in the treatments receiving terrigenic inputs in the dominant phytoplankton species over time. Initially, the phytoplankton community was dominated by Chlorophyta species such as Chlorella sp., Selenastrum minutum, and Crucigenia quadrata. However, as the experiment progressed, there was a transition towards a Cyanobacteria-dominated state (i.e. Pseudanabaena sp., Microcystis wesenbergii, Aphanizomenon flosaquae, Oscillatoria tenuis, etc.). Notably, different terrigenic treatments exhibited a co-dominance of Chlorella sp., Pseudanabaena sp., and Microcystis wesenbergii at different experiment stages.
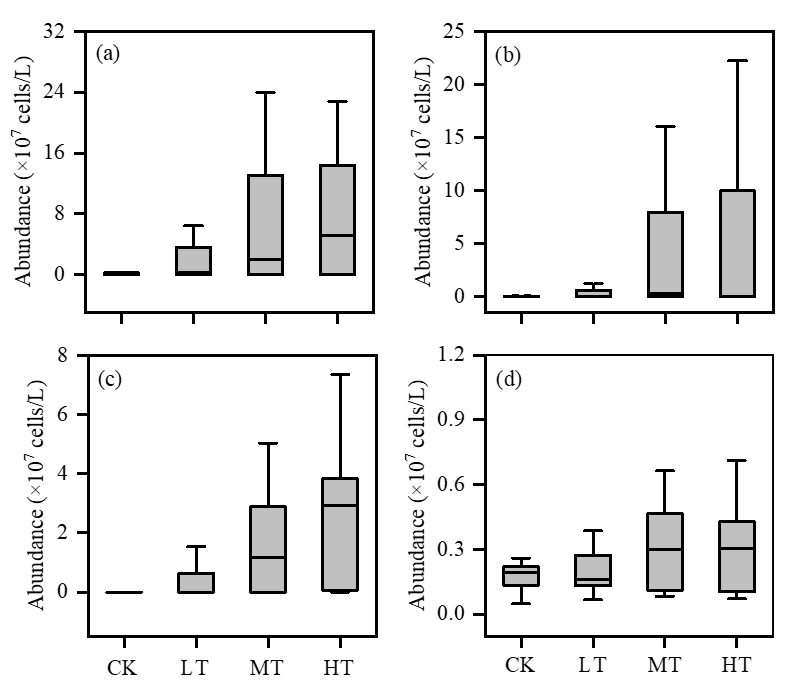
The abundance of the four dominant phytoplankton species, namely Pseudanabaena sp., Microcystis wesenbergii, Aphanizomenon flosaquae, and Chlorella sp., followed a similar increasing pattern across the different terrigenic treatments in the experiment (Figure 5). In the CK treatment, the mean abundance of Pseudanabaena sp., Microcystis wesenbergii, and Chlorella sp. was recorded as (0.11 ± 0.10) × 107, (0.09 ± 0.21) × 107, and (0.17 ± 0.06) × 107 cells/L, respectively. However, in the MT treatment, their abundance increased significantly to (11.16 ± 16.90) × 107, (5.09 ± 7.65) × 107, and (0.39 ± 0.38) × 107 cells/L, respectively (Figure 5a, b, d). Similarly, the abundance of Aphanizomenon flosaquae increased from (0.05 ± 0.11) × 107 cells/L in the CK treatment to (3.10 ± 3.54) × 107 cells/L in the HT treatment (Figure 5c). The abundance of all four dominant species was significantly (P < 0.05) higher in the MT and HT treatments compared to the CK treatment.
_*pseudanabaena*_sp.__(b)_*microcystis_wesenbergii*__(c)_*aphanizomeno.png)
The experiment revealed a decreasing trend in phytoplankton diversity indices across the three terrigenic treatments (Figure 6). The mean values of Pielou’s evenness and Shannon-Wiener diversity increased from 0.65 ± 0.15 and 1.47 ± 0.44 in the CK treatment to 0.87 ± 0.07 and 1.83 ± 0.20 in the LT treatment, respectively. Notably, the phytoplankton diversity indices were significantly (P < 0.05) higher in the LT treatment compared to the CK treatment.
_and_shannon-wiener_diversity_(b)_in_different_terrigen.png)
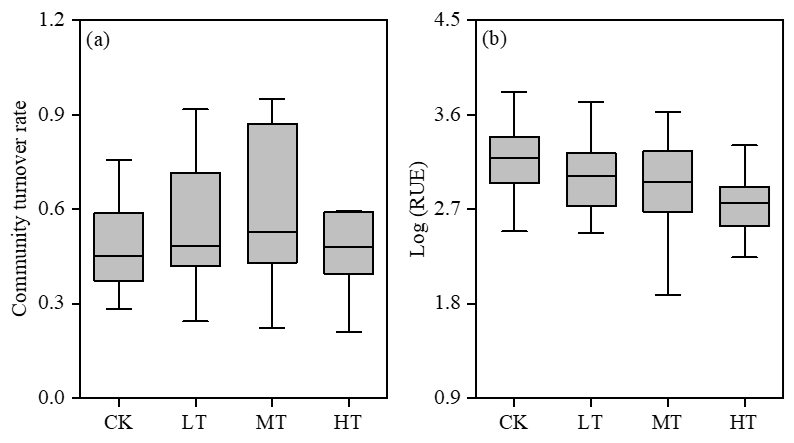
The experiment revealed that the terrigenic input impacted the phytoplankton community’s turnover rate (Figure 7a). The mean community turnover rate increased from 0.49 ± 0.15 in the CK treatment to 0.61 ± 0.25 in the MT treatment. Conversely, phytoplankton resource use efficiency displayed a significant (P < 0.05) negative response to increased terrigenic input (Figure 7b). The mean resource use efficiency decreased from 3.10 ± 0.44 in the CK treatment to 2.66 ± 0.47 in the HT treatment.
_and_phytoplankton_resource_use_efficiency_(b)_in.png)
The resistance of phytoplankton communities in the three terrigenic treatments was observed to have higher values than the CK treatment (Figure 8). Throughout the entire experiment, the LT treatment displayed frequent temporal fluctuations and remained closest to the CK treatment in terms of resistance (Figure 8a). However, its performance was significantly (P < 0.05) higher than the CK treatment by the end of the experiment. On the other hand, the MT and HT treatments initially approached the CK treatment but had significantly (P < 0.05) higher resistance values compared to the CK treatment after the first week (Figure 8b, c). Among the three treatments, only the HT treatment exhibited a significant (P < 0.05) positive correlation with the stability of phytoplankton community.

Discussion
It is widely acknowledged that rainfall events can potentially reduce phytoplankton diversity indices, resulting in a more homogenous species composition and increased ecosystem vulnerability.28–31 Previous studies have indicated that intense rainfall indirectly promotes phytoplankton growth and triggers cyanobacterial blooms by enhancing nutrient inputs into the water column.32 Expanding on this understanding, our research demonstrates that simulated terrigenic inputs significantly impact phytoplankton communities’ composition, abundance, and diversity indices. Consistent findings from our modeling experiments have established a strong association between heavy rainfall events and changes in the structure (e.g. species composition, abundance, diversity indices, and community turnover) and functioning (e.g. resource use efficiency) of phytoplankton communities within lake ecosystems.
The species richness of phytoplankton plays a crucial role in shaping the structure and functioning of aquatic environments.33 In our experiment, a comprehensive analysis identified 90 phytoplankton species belonging to 18 genera and 7 phyla. The findings indicated a declining trend in species richness across the four terrigenic treatments, suggesting that high terrigenic input can diminish its impact on the phytoplankton community, reducing species diversity.34 Conversely, the abundance of phytoplankton exhibited an increasing trend across the four terrigenic treatments, indicating that high terrigenic input can stimulate phytoplankton reproduction, consistent with Vizzo et al.35
The dominant species within a community play a crucial role in shaping energy transfer, material cycling, and the overall trajectory of community succession.36 In the context of lake ecosystems, heavy rainfall events leading to increased nutrient concentrations can disrupt the succession patterns of dominant phytoplankton species.37 In our study, we observed distinct taxon-specific trajectories in response to terrigenic input, with Chlorophyta and cyanobacteria dominating in the CK treatment and cyanobacteria exhibiting a significant increase in abundance under the terrigenic treatments. Furthermore, our observations revealed that higher nutrient concentrations correlated with a higher proportion of cyanobacteria, indicating that nutrient input from heavy rainfall raises the risk of cyanobacterial outbreaks in the lake. These findings align with the findings of Richardson,38 who also concluded that elevated nutrient loads contribute to an increase in cyanobacterial abundance.
The phytoplankton diversity index serves as a valuable metric for assessing the structure of phytoplankton communities, with higher diversity indicating a more intricate community structure, as well as greater stability and resilience.39 This study observed a decline in phytoplankton diversity in the terrigenic treatments as nutrient loads increased. This decline can be attributed to the dominance of a single species resulting from the terrigenic input, which simplifies the community structure and reduces its stability. These findings suggest that heavy rainfall events augment the input of terrestrial sources’ nutrients, compromising the lake ecosystem’s stability. Similar conclusions were drawn by Ciglenečki et al.,40 who reported a negative correlation between nutrient concentrations and phytoplankton diversity. This highlights the significant impact of higher terrigenic input on phytoplankton diversity, ultimately disrupting the structure of the lake’s phytoplankton community.41
Our findings indicate a strong association between high terrigenic input and increased phytoplankton community turnover throughout the experiment. This phenomenon can be attributed to elevated nitrogen and phosphorus concentrations resulting from terrestrial nutrient inputs, creating a favorable environment for phytoplankton colonization.42 A similar study conducted by Villafañe et al.43 yielded similar results, further supporting our findings. Community turnover can profoundly impact ecosystem functioning, especially in disturbance regime-dependent ecosystems.44 Resource use efficiency (RUE) of phytoplankton, a crucial ecological concept reflecting their ability to capture resources, also plays a key role in ecosystem functioning.45 Interestingly, our study reveals a decline in the efficiency of phytoplankton resource use with increasing terrigenic input, possibly due to the toxic effects of the high terrigenic input on the phytoplankton community.46,47
Previous research has established that ecosystems with high species diversity exhibit greater long-term stability.26 However, the impact of terrigenic input as a disturbance factor on phytoplankton community structure has received limited attention. Our study observed an overall increase in total phytoplankton abundance across all terrigenic treatments compared to the control group, suggesting that higher terrigenic inputs may contribute to enhanced phytoplankton abundance. Resistance, which refers to the extent of change in phytoplankton abundance in response to disturbances, often indicates ecological stability.48 In this study, all treatments displayed rapid responses to the terrigenic input. Notably, the low and medium terrigenic input systems showed signs of recovery by the end of the experiment, supporting the notion that ecosystems with higher diversity tend to be more stable.49 However, the high terrigenic input system did not exhibit recovery and had lower genus richness than the low and medium terrigenic input systems. Despite this, it demonstrated higher resistance in our experiment. The terrigenic treatments were dominated by advantageous cyanobacteria, which have a better chance of survival under high nutrient conditions than non-cyanobacterial groups. This finding aligns with the results of Filiz et al.,50 who found that nutrient loading had the strongest impact on the phytoplankton community.
Conclusion
This study employed in situ models to investigate the impact of significant rainfall events on the phytoplankton community structure of Lake Changhu. The results revealed that heavy rainfall events led to an increase in nutrient concentration, causing a shift in the original phytoplankton composition towards a cyanobacteria-dominated community. This shift was positively correlated with rainfall intensity, and intense rainfall episodes also decreased the phytoplankton diversity index and community conversion rate. These findings highlight the potential occurrence of cyanobacterial blooms in Lake Changhu due to heavy rainfall events and emphasize the need for stronger measures to prevent bloom outbreaks during the summer months when the lake is particularly susceptible to heavy rainfall influences.
Acknowledgments
This research was financially supported by the National Natural Science Foundation of China (U21A2039, 41901135). We thank Jingzhou Meteorological Bureau for providing the meteorological data for this manuscript.
Authors’ Contribution according to CRediT
Conceptualization: Yefei Zhang (Equal), Jun R. Yang (Equal). Methodology: Yefei Zhang (Equal), Tao Li (Equal), Lu Zhang (Equal), Qin Hu (Equal). Formal Analysis: Yefei Zhang (Equal), Tao Li (Equal), Lu Zhang (Equal), Qin Hu (Equal). Investigation: Yefei Zhang (Equal), Tao Li (Equal), Lu Zhang (Equal), Qin Hu (Equal). Writing – original draft: Yefei Zhang (Equal), Jun R. Yang (Equal). Writing – review & editing: Yefei Zhang (Equal), Jun R. Yang (Equal). Resources: Zhangyong Liu (Equal), Yi Chai (Equal). Supervision: Zhangyong Liu (Equal), Jianqiang Zhu (Equal), Jun R. Yang (Equal). Funding acquisition: Jianqiang Zhu (Equal), Jun R. Yang (Equal).